|
AUGUSTOWSKO-SUWALSKIE TOWARZYSTWO NAUKOWE
Proszę chwilę zaczekać, ładuję stronę... |
|
AUGUSTOWSKO-SUWALSKIE TOWARZYSTWO NAUKOWE
Proszę chwilę zaczekać, ładuję stronę... |
|
Maciej Wódkiewicz
Dynamika populacji kukuczki kapturkowatej Neottianthe cucullata (L.) Schlechter w Wigierskim Parku Narodowym
Kukuczka kapturkowata Neottianthe cucullata (L.) Schlechter jest niewielką rośliną posiadającą jeden lub dwa eliptyczne liście, rozpościerające się płasko na ziemi. Podczas kwitnienia rośliny osiągają wysokość 15 (30) cm. Kwiatostan składa się z kilku do kilkunastu różowobiałych drobnych kwiatów. Kukuczka rośnie w mszystych borach sosnowych i sosnowo-świerkowych w pobliżu zbiorników wodnych. Jest to jedyny europejski przedstawiciel azjatyckiego rodzaju Neottianthe. Gatunek ten osiąga w Polsce południowo-zachodni kres zasięgu (Filipek 1956; Hulten, Fries 1986). W Polsce kukuczka występuje na niewielkiej liczbie stanowisk (Zając, Zając 2001). Jedno z nich jest położone na terenie Wigierskiego Parku Narodowego. Podobnie jak na Białorusi, Litwie, Ukrainie i w obwodzie kaliningradzkim jest ona uważana za gatunek zagrożony wyginięciem – EN (Kruszelnicki 2001). Jako główne przyczyny zagrożenia podaje się wyrąb lasów i procesy naturalnej sukcesji (Kruszelnicki 2001; Piękoś-Mirkowa, Mirek 2003). Niniejsza praca przedstawia wyniki pierwszej serii z zaplanowanych na wiele lat badań nad dynamiką populacji tego gatunku w Wigierskim Parku Narodowym. Celem pracy była odpowiedź na następujące pytania: Czy liczebność lokalnej populacji się zmniejsza? Czy w lokalnej populacji zachodzi rekrutacja nowych osobników? Jaki jest los nowo pojawiających się osobników? Jaka jest długość życia osobnika? Przez ile lat osobniki przystępują do reprodukcji?
Metody
W latach 1998–2003 w porze kwitnienia kukuczki na początku sierpnia kartowałem wszystkie osobniki na stałej powierzchni badawczej, założonej w 1998 roku na terenie zajmowanym przez populację tego gatunku w Wigierskim Parku Narodowym. Powierzchnia miała kształt prostokąta 5 m x 8 m (40 m2). Dla każdego osobnika odnotowałem liczbę liści i obecność kwiatostanu. Określiłem fazę rozwojową każdego osobnika na podstawie obecności kwiatostanu oraz obecności danego osobnika w roku poprzednim. Osobniki z pędem kwiatowym zakwalifikowałem jako osobniki generatywne. Osobniki bez pędu kwiatowego zakwalifikowałem jako wegetatywne, jeśli odnotowałem je w latach ubiegłych, natomiast jako osobniki juwenilne, jeśli nie odnotowałem ich wcześniej na badanej powierzchni. Osobniki, które nie pojawiły się w danym roku, a były odnotowane wcześniej i w latach następnych, określiłem jako utajone. W 1998 roku osobniki nieposiadające pędu kwiatowego zostały zakwalifikowane do grupy „nieokreślone”, ze względu na brak informacji o wcześniejszym ich występowaniu na badanej powierzchni.
Wyniki
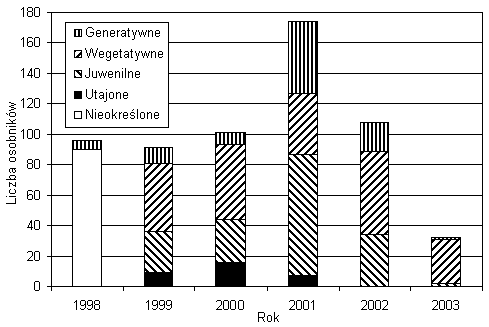
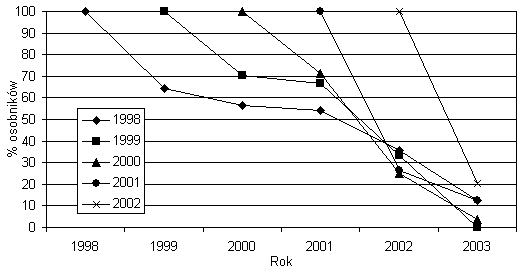
Średnia liczebność populacji w poszczególnych latach wynosiła około stu osobników. Fluktuacje rzędu 70 proc. w porównaniu ze średnią dla całego okresu badań liczebnością odnotowałem w 2001 i 2003 roku (rys. 1). Okres półtrwania kohort z lat 1999–2002 wynosił odpowiednio 2,5; 1,5; 0,7 i 0,6 roku. Największe straty w liczbie osobników ze wszystkich kohort odnotowałem pomiędzy 2001 i 2002 rokiem (rys. 2). Podczas sześciu lat obserwacji nie odnotowałem tendencji spadkowej w liczebności populacji.
Rys. 1. Liczba osobników i struktura populacji w kolejnych latach
W latach 1999–2003 obserwowałem pojawienie się nowych osobników w populacji. W 1998 roku nie byłem w stanie rozróżnić osobników pomiędzy wegetatywnymi a juwenilnymi ze względu na brak informacji o obecności konkretnych osobników na powierzchni w latach poprzedzających. Największą rekrutację odnotowałem w roku 2001, najmniejszą w 2003 (rys. 1). Nie licząc tych dwóch lat, przeciętna rekrutacja kształtowała się na poziomie 30 osobników rocznie, co stanowiło około 30 proc. osobników odnotowanych w danym roku. Każdego roku (oprócz 1998) odnotowałem od 30 do 60 osobników wegetatywnych. W latach 1999, 2000, 2002 stanowiły one około 50 proc. osobników, w roku 2001 – 24 proc., a w 2003 – 91 proc. wszystkich osobników. W poszczególnych latach odnotowałem od jednego (rok 2003) do blisko 50 (2001) osobników generatywnych. Stanowiły one od kilku do blisko 30 proc. populacji.
Rys. 2. Krzywe przeżywalności kohort 1999–2002 i
różnowiekowej populacji osobników odnotowanych
Rys. 3. Losy nowych osobników (kohorty 1999–2002)
Spośród kohort 1999–2002 czterdzieści sześć proc. nowych osobników miało jeden liść, a 49 proc. osobników dwa liście (rys. 3). Pozostałe 5 proc. stanowiły osobniki z dwoma liśćmi kwitnące w pierwszym roku. Do kwitnienia w kolejnych latach przystępuje stosunkowo więcej osobników posiadających w pierwszym roku dwa liście niż posiadających w pierwszym roku życia jeden liść. Osiemdziesiąt trzy proc. osobników kwitło tylko raz w trakcie okresu badań, a 17 proc. dwa razy. Wszystkie osobniki kwitnące dwa razy miały w pierwszym roku dwa liście.
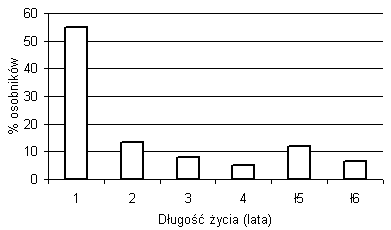
Ponad 50 proc. nowych osobników żyje tylko przez jeden rok, nie pojawiając się w latach następnych (rys. 4). Osobniki dwu-, trzy- i czteroletnie stanowią odpowiednio od kilkunastu do kilku procent populacji. Pewna część osobników ma dłuższy okres życia. Dwanaście osobników odnotowanych w 1998 roku było obecnych na badanej powierzchni w roku 2003.
Rys. 4. Długość życia osobników
Dyskusja
Fluktuacje liczby osobników w populacji mogły być, przynajmniej częściowo, powodowane warunkami pogodowymi. Można przypuszczać, że ciepłe i wilgotne okresy letnie oraz ciepłe zimy sprzyjają wzrostowi liczebności populacji, natomiast suche lata i mroźne zimy wpływają na zmniejszenie się liczebności populacji. Powodem zmniejszenia się liczebności populacji w 2002 i 2003 roku mogła być przede wszystkim mroźna zima (tab. 1). Czas półtrwania kohort 1999–2001 oraz populacji osobników odnotowanych w 1998 roku przypada pomiędzy 2001 a 2002 rokiem. Przyczyną tego może być mroźna zima 2001/2002. Niekorzystne warunki pogodowe mogą mieć negatywny wpływ na długość półtrwania kohort kukuczki. Aby zweryfikować wpływ warunków pogodowych na długość życia osobników i liczebność populacji, niezbędna jest dłuższa seria badań. Obserwowane przeze mnie fluktuacje są dużo mniejsze niż podawane przez innych autorów (Filipek 1956; Wiśniewski 1976). Może to być związane z tym, że za miarę wielkości populacji przyjęli oni jedynie liczbę osobników kwitnących, nie biorąc pod uwagę osobników wegetatywnych.
Tabela 1. Warunki pogodowe w kolejnych latach
Pojawianie się każdego roku nowych osobników w populacji świadczy o tym, że zachodzi rekrutacja i populacja stale się odnawia. Dopływ około 30 proc. osobników juwenilnych każdego roku oraz średni okres półtrwania kohort wynoszący około dwóch lat, świadczy o szybkim tempie zastępowania starych osobników przez nowe. Obecność młodych osobników gwarantuje możliwość trwania populacji tego gatunku na badanym terenie. Wysoka rekrutacja w 2001 roku mogła być powodowana sprzyjającymi warunkami pogodowymi. Przyczyną niskiej rekrutacji w roku 2003 była najprawdopodobniej mroźna zima (tab. 1). Obecność osobników generatywnych w każdym roku badań gwarantowała stały dopływ nasion i możliwość rekrutacji. Udział osobników kwitnących w populacji nie był stały. Oszacowanie liczebności populacji na podstawie liczby osobników kwitnących nie jest dokładne. Duży procent osobników żyjących tylko przez jeden rok świadczy o wysokim prawdopodobieństwie wyginięcia w początkowym okresie. Niewielki procent osobników, które przeżyją ten niekorzystny okres, jest w stanie trwać dłużej. Konkurencyjność osobników, które w pierwszym roku pojawiły się z dwoma liśćmi, w stosunku do osobników, które pojawiły się w pierwszym roku z tylko jednym liściem, może być powodowana większą ilością zasobów zgromadzonych w fazie podziemnej. Najprawdopodobniej umożliwia to wytworzenie większego aparatu asymilacyjnego i produkcję większej ilości substancji zapasowych, co pozwala przeżyć roślinom przez dłuższy czas. Większa ilość zgromadzonych substancji zapasowych może zwiększyć również prawdopodobieństwo zakwitnięcia. Koszty reprodukcji są najprawdopodobniej dość wysokie, dlatego tylko niewielka część osobników przystępuje do kwitnienia więcej niż raz. Największym zagrożeniem dla kukuczki kapturkowatej jest gospodarka leśna związana z pozyskiwaniem drewna. Można mieć nadzieję, że populacja tego gatunku, znajdująca się na terenie Wigierskiego Parku Narodowego, będzie miała szansę przetrwać dzięki zapewnianej przez park ochronie.
Podsumowanie
Badaniami objęto populację kukuczki kapturkowatej (Neottianthe cucullata) w Wigierskim Parku Narodowym. W latach 1998–2003 odnotowywano rocznie około 100 osobników. Wydaje się, że mroźne zimy mają wpływ na zmniejszenie liczebności populacji w danym roku zarówno poprzez zmniejszenie rekrutacji, jak i zwiększenie śmiertelności. Każdego roku obserwowano kwitnienie oraz pojawianie się nowych osobników. Osobniki juwenilne stanowiły około 30 proc. wszystkich osobników odnotowanych w danym roku. Ponad połowa osobników żyje jeden rok. Wysoki procent nowych osobników oraz krótki czas ich życia świadczą o szybkim tempie zastępowania starych osobników nowymi. Osobniki pojawiające się w pierwszym roku z dwoma liśćmi stanowiły około połowę nowych osobników. Więcej osobników kwitło spośród pojawiających się w pierwszym roku z dwoma liśćmi niż pojawiających się w pierwszym roku z jednym liściem. Podczas sześcioletniego okresu badań większość osobników przystąpiła do kwitnienia tylko jeden raz.
Serdecznie dziękuję dyrekcji Wigierskiego Parku Narodowego za możliwość prowadzenia badań oraz Marcie Badurek za pomoc w gromadzeniu danych w terenie. Projekt był finansowany z funduszy KBN na badania własne dla Wydziału Biologii UW, BW nr 1521/55 oraz Komitet Badań Naukowych – grant nr 3P04G 073 22.
Abstract
Population dynamics of Neottianthe cucullata (L.) Schlechter in Wigry National Park A population of Neottianthe cucullata has been studied in Wigry National Park. It is one of a few populations of this endangered species (category EN of Polish Red Data Book) existing in Poland. During six years (1998–2003) about 100 individuals were recorded each year. Cold winters probably cause decrease in number of individuals in the local population through higher mortality and reduced recruitment. Each year flowering individuals and new recruits were observed. New individuals comprised about 30% of plants observed each year of this study. Over 50% of individuals live for only one year. High fraction of new individuals and their short life span indicate a quick turnover of individuals in this population. Individuals appearing for their first time with two leaves made up about half of new recruits. There were more flowering individuals among those appearing for the first time with two leaves in comparison with individuals appearing for the first time with only one leaf. Most of flowering individuals flowered only once during this study. Those that flowered twice appeared for the first time above ground with two leaves.
Literatura
Filipek M., 1956: Neottianthe cucullata (L.) Schlechter w Puszczy Augustowskiej. Spraw. Poznańskiego Tow. Przyjaciół Nauk, III-IV, 48–53. Hulten E., Fries M., 1986: Atlas of North European Vascular Plants. T. 1. Königstein, Koeltz Scientific Books, t. 1, 3:280. Kruszelnicki J., 2001: Neottianthe cucullata. W: Kaźmierczakowa R., Zarzycki K. (red.): Polska czerwona księga roślin. Kraków, Polska Akademia Nauk, Instytut Botaniki im. W. Szafera, Instytut Ochrony Przyrody, s. 545–546. Piękoś-Mirkowa H., Mirek Z., 2003: Flora Polski. Atlas roślin chronionych. Warszawa, „Multico” Oficyna Wyd., s. 314–315. Wiśniewski N., 1976: Zum Verhalten von Neottianthe cucullata (L.) Schlt. An der Westgrenze ihrer Verbreitung in Europa. Berlin, In Feddes Repertorium, Band 87, Heft 9–10:587–609. Zając A., Zając M. (red.), 2001: Atlas rozmieszczenia roślin naczyniowych w Polsce. Kraków, Inst. Bot. UJ.
|