|
AUGUSTOWSKO-SUWALSKIE TOWARZYSTWO NAUKOWE
Proszę chwilę zaczekać, ładuję stronę... |
|
AUGUSTOWSKO-SUWALSKIE TOWARZYSTWO NAUKOWE
Proszę chwilę zaczekać, ładuję stronę... |
|
Powszechnie uważa się, że gatunki o dużych kwiatostanach są atrakcyjniejsze dla zwierząt zapylających kwiaty, co przekłada się na zwiększony sukces reprodukcyjny tych roślin (Jong de, Klinkhamer 1994). Zagadnienie to było studiowane teoretycznie (Ohashi, Yahara 1999) i choć uzyskane wyniki w ogólności potwierdzają przypuszczenia o większej atrakcyjności dużych kwiatostanów, to wskazują również, że duże znaczenie może mieć odległość dzieląca poszczególne rośliny, a być może także części kwiatostanu. Gatunkami, które doskonale nadają się do badania takich zależności, wydają się rośliny z rodziny baldaszkowatych (Apiaceae = Umbelliferae). W ich przypadku już dawniej badacze uzależniali stopień obcopylności roślin od struktury i wielkości kwiatostanu (Bell 1971). Teoretyczne modele przewidują, że kwiatostany o luźnej strukturze wymuszają na owadach przemieszczanie się pomiędzy baldachami, zwiększając dzięki temu stopień allogamii, natomiast kwiatostany zwarte powinny powodować wydłużenie wizyt w obrębie jednego kwiatostanu (Bell, Lindsey 1978). Spośród europejskich baldaszkowatych doskonałym modelem do tego typu badań jest Heracleum sphondylium, którego kwiatostany mogą znacznie różnić się wielkością oraz – w zależności od podgatunku – stopniem zwartości baldachów.
Materiał i metody
Aktywność owadów na kwiatostanach dwóch podgatunków H. sphondylium: nominatywnego (H. sphondylium subsp. sphondylium) i syberyjskiego (H. sphondylium subsp. sibiricum), określałem na podstawie analizy filmu nakręconego kamerą wideo oraz odłowów owadów odwiedzających kwiaty. Wybrane podgatunki różnią się budową morfologiczną kwiatów i kwiatostanów. Pierwszy z nich ma białe kwiaty, z których większość charakteryzuje się symetrią dwuboczną. Baldachy podgatunku nominatywnego są gęste i tworzą jedną powierzchnię, są też zwykle większe od kwiatostanów podgatunku syberyjskiego. Kwiaty podgatunku syberyjskiego są żółtozielone i promieniste, zaś jego baldachy mają luźną strukturę (Zych 2002). Obserwacje przeprowadziłem w sezonach 2000 i 2002 na terenie Wigierskiego Parku Narodowego (tab. 1), jednym z niewielu miejsc w Polsce, w którym oba te taksony występują sympatrycznie. Odłowów i nagrań dokonywałem w czasie przypadającym na szczyt kwitnienia (lipiec), na kwiatach w obu fazach płciowych w godzinnych interwałach między ósmą a dziewiętnastą. Analizowałem liczbę wizyt owadów z różnych rzędów (Coleoptera, Diptera, Heteroptera, Hymenoptera oraz Lepidoptera), uzależniając ją od takich parametrów, jak: rok badań, podgatunek H. sphondylium, faza płciowa kwiatów, pora dnia, wielkość kwiatostanu (mierzona liczbą baldaszków) czy warunki pogodowe (siła wiatru, zachmurzenie). Ponadto na podstawie nagrań wideo badałem zachowanie owadów na kwiatostanach, w szczególności notując dystans pokonywany przez poszczególne gatunki na kwiatostanach obu podgatunków H. sphondylium.
Tabela 1. Opis stanowisk na terenie Wigierskiego Parku Narodowego, gdzie przeprowadzono badania zapylaczy dwóch podgatunków H. sphondylium
Wyniki i dyskusja
W przypadku H. sphondylium wielkość kwiatostanów nie ma wpływu na częstość odwiedzin zapylaczy (muchówek i chrząszczy) (tab. 2). Możliwe, co sugerują modele Ohashiego i Yahary (1999), że w przypadku badanych roślin, rosnących w zwartej populacji, znacznie ważniejsze dla zapylaczy są odległości pomiędzy poszczególnymi kwiatostanami niż wielkość pojedynczego baldachu. Badania wykazały, iż czynnikami wpływającymi na częstość wizyt są natomiast: struktura kwiatostanu (podgatunek), pora dnia, rok badań oraz faza płciowa kwiatostanów. Spośród parametrów pogodowych większe znaczenie ma siła wiatru (tab. 2).
Tabela 2. Uproszczone wyniki analizy komponentów wariancyjnych (efekty proste). Testowano wpływ następujących czynników: badana populacja, sezon badań (rok), faza płciowa kwiatostanów (płeć), pora dnia, zachmurzenie, siła wiatru oraz wielkość kwiatostanu mierzona liczbą baldaszków (kwiatostan) na liczbę wizyt owadów poszczególnych rzędów na kwiatach dwóch podgatunków H. sphondylium. Do analizy użyto danych dla sezonów 2000 i 2002, blokując wyniki w grupy odpowiadające czterem porom dnia: rankowi, południowi, popołudniowi i wieczorowi (N=192 obserwacje). Prawdopodobieństwo oznaczone odpowiednio: *** p<0,005, ** p<0,01, * p<0,05, puste miejsce oznacza brak istotnego wpływu badanego czynnika, Σ – całkowita liczba wizyt
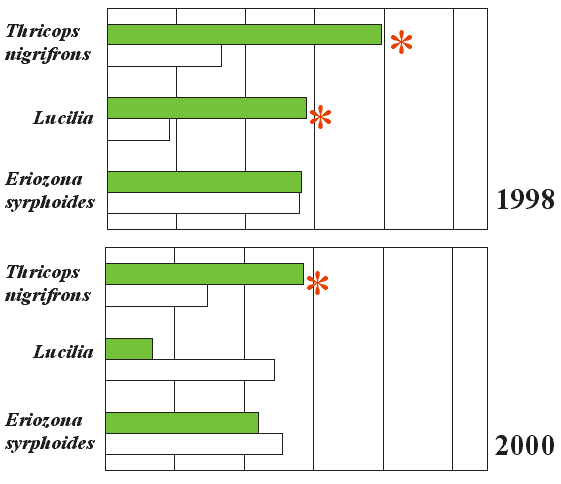
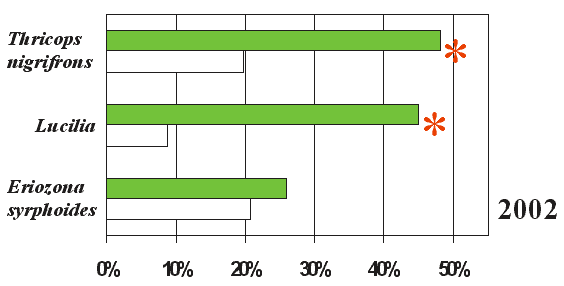
We wcześniejszych badaniach różnych gatunków baldaszkowatych postulowano już związek atrakcyjności kwiatostanów (mierzonej stopniem zawiązywania nasion) z ich budową (m.in. gęstością baldachów). Zależność taką odkryto dla dwóch gatunków z rodziny Apiaceae (Thaspium barbinode i Zizia trifoliata) badanych przez Lindsey (1984). Jeżeli zważyć, że sukces reprodukcyjny baldaszkowatych nie jest zwykle limitowany przez dostępność zapylaczy (Bell, Lindsey 1978), wyjaśnieniem tego zjawiska może być wpływ jakości i ilości pyłku na jakość nasion. Wiadomo, że owoce zawiązane po zapyleniu dokonanym większą liczbą ziaren pyłku są żywotniejsze (Wilcock, Neiland 2002). Prawdopodobnie istnieje nacisk selekcyjny na utrzymanie dużych kwiatostanów, które, chociaż kosztowne energetycznie (Harder, Barret 1995), dzięki swej atrakcyjności dla wielu owadów, a co za tym idzie masowemu przepływowi pyłku, zapewniają bardziej heterozygotyczne potomstwo (Bell, Lindsey 1978) oraz większy męski sukces reprodukcyjny. Budowa baldachów może także tłumaczyć zachowanie owadów. Wydaje się, że luźniejsze kwiatostany, w których poszczególne baldaszki nie są połączone w jedną powierzchnię, powinny wywoływać zwiększony ruch pomiędzy roślinami (Bell, Lindsey 1978) i zmniejszać liczbę baldaszków odwiedzonych za jednym razem. Baldachy zwarte stymulują większą aktywność w obrębie kwiatostanu. U niektórych gatunków, np. amerykańskich przedstawicieli rodzajów Thaspium i Zizia, obserwuje się zmianę gęstości kwiatostanów w przeciągu okresu kwitnienia – kwiatostany w fazie żeńskiej są bardziej zwarte, baldachy w fazie męskiej zaś bardziej rozpierzchłe (Lindsey, Bell 1985). W pierwszym przypadku służy to prawdopodobnie zwiększeniu liczby zapyleń, w drugim natomiast wzmaga transfer pyłku. W przypadku badanych podgatunków barszczu zwyczajnego nie zauważono takich zmian w morfologii kwiatostanów w trakcie kwitnienia, jednak oba studiowane taksony różniły się pod tym względem od siebie. W myśl opisanych wcześniej założeń, owady powinny odwiedzać mniej baldaszków na kwiatostanach podgatunku o luźniejszych baldachach – w tym wypadku barszczu syberyjskiego. Analiza aktywności owadów przyniosła jednak zgoła odwrotne wyniki – w przypadku większości owadów nie zanotowano istotnych różnic w liczbie baldaszków odwiedzanych na kwiatostanach jednego i drugiego taksonu, natomiast w przypadku owadów, dla których takie różnice istniały, układały się one odwrotnie od teoretycznych założeń. Główne zapylacze tej rośliny: muchówki Thricops nigrifrons i Lucilia spp. (Zych 2002), odwiedzały mniej baldaszków na bardziej zwartych kwiatostanach subsp. sphondylium, dla syrfidy Eriozona syrphoides nie zanotowano żadnych różnic (rys. 1). Możliwe, że na wyniki wpływają interakcje wspomnianych muchówek z innymi owadami. Na kwiatach podgatunku nominatywnego masowo występują niewielkie muchówki z rodziny wońkowatych (Sepsidae), które – jak obserwowano – dość często odganiały od zajmowanych przez siebie baldaszków inne, nawet stosunkowo duże muchówki, skracając przez to ich wizyty. Osobniki Lucilia spp. i T. nigrifrons są jednymi z częstszych owadów na kwiatach H. sphondylium subsp. sphondylium, więc to właśnie one są najczęstszymi obiektami ataków Sepsidae. Dodatkowo kwiatostany barszczu są patrolowane przez drapieżne błonkówki poszukujące ofiar. Niewykluczone, że w przypadku białego, prawie jednolitego tła, jakie dają kwiatostany subsp. sphondylium, owady żerujące na nich są znacznie lepiej widoczne niż na zielonkawożółtych, ażurowych kwiatostanach subsp. sibiricum, a to także wpływa na skracanie wizyt na tych pierwszych. Problem zachowania owadów na kwiatach oraz wpływu budowy kwiatostanów na sukces reprodukcyjny roślin, choć bardzo interesujący i ważny z punktu widzenia strategii reprodukcyjnych, nie był prawie studiowany w przypadku baldaszkowatych i zdecydowanie wymaga dalszych badań.
Rys. 1. Zachowanie najważniejszych zapylaczy na kwiatostanach w fazie męskiej dwóch podgatunków H. sphondylium: subsp. sibiricum (kolor zielony) oraz subsp. sphondylium (kolor biały) w sezonach 1998, 2000 i 2002. Dystans pokonywany przez owady pokazano jako średnią liczbę baldaszków odwiedzonych w stosunku do średniej liczby baldaszków w kwiatostanie (x 100 proc.). Gwiazdką oznaczono wyniki istotne statystycznie przy p<0,05 (test T-studenta lub Z, wykonany dla danych surowych)
Podsumowanie
Wytwarzanie dużych kwiatostanów jest uważane za sposób przywabiania znacznej liczby potencjalnych zapylaczy. Badania przeprowadzone w Wigierskim Parku Narodowym na dwóch podgatunkach H. sphondylium (Apiaceae) wykazały jednak, że czynnik ten nie tłumaczy atrakcyjności poszczególnych roślin. Znacznie większe znaczenie mają inne cechy kwiatostanów – pora dnia, niektóre parametry pogodowe (głównie siła wiatru) i inne czynniki. Obserwacje wskazują także, że interakcje z innymi owadami odwiedzającymi kwiaty H. sphondylium mogą skracać wizyty znaczących zapylaczy.
Literatura
Bell C. R., 1971: Breeding systems and floral biology of the Umbelliferae or evidence for specialization in unspecialized flowers. W: Heywood V. H. (red.): The Biology and Chemistry of the Umbelliferae. London, Academic Press, s. 93–107. Bell C. R., Lindsey A. H., 1978: The umbel as a reproductive unit in the Apiaceae. W: Actes du 2eme Symposium International sur les Ombelliferes. Contributions pluridisciplinaires a la Systematique. France, Perpignan, s. 739–747. Harder L. D., Barrett S. C. H., 1995: Mating costs of large floral displays in hermaphrodite plants. Nature, 373:512–515. Jong T. J. de, Klinkhamer P. G. L., 1994: Plant size and reproductive success through female and male function. Journal of Ecology, 82:399–402. Lindsey A. H., 1984: Reproductive biology of Apiaceae. 1: Floral visitors to Thaspium and Zizia and their importance in pollination. American Journal of Botany, 71:375–387. Lindsey A. H., Bell C. R., 1985: Reproductive biology of Apiaceae. 2: Cryptic specialization and floral evolution in Thaspium and Zizia. American Journal of Botany, 72:231–247. Ohashi K., Yahara T., 1999: How long to stay on, and how often to visit a flowering plant? – a model for foraging strategy when floral displays vary in size. Oikos, 86:386–392. Wilcock C., Neiland R., 2002: Pollination failure in plants: why it happens and when matters. Trends in Plant Sciences, 7:270–277. Zych M., 2002: Pollination biology of Heracleum sphondylium L. (Apiaceae). The advantages of being white and compact. Acta Societatis Botanicorum Poloniae, 71:163–170.
|