|
AUGUSTOWSKO-SUWALSKIE TOWARZYSTWO NAUKOWE
Proszę chwilę zaczekać, ładuję stronę... |
|
AUGUSTOWSKO-SUWALSKIE TOWARZYSTWO NAUKOWE
Proszę chwilę zaczekać, ładuję stronę... |
|
Ewolucja systemów zapylania na przykładzie europejskich przedstawicieli rodzaju Heracleum L. (Apiaceae)
Rodzaj Heracleum L., należący do rodziny baldaszkowatych (Apiaceae), jest w Europie reprezentowany przez dziewięć gatunków (Brummitt 1968). Stosunki pokrewieństwa pomiędzy nimi nie są jak do tej pory zbadane. Spośród europejskich barszczy siedem to gatunki endemiczne rejonów górskich, jak np. barszcz austriacki (H. austriacum), występujący w austriackich Alpach, oraz jeden gatunek inwazyjny – zawleczony z Azji barszcz kaukaski (H. mantegazzianum). Dziewiąty gatunek to pospolicie występujący, polimorficzny barszcz zwyczajny (H. sphondylium), będący częstym gatunkiem łąkowym, zaroślowym i ruderalnym w wielu krajach europejskich (Sheppard 1991).
Heracleum sphondylium – model w badaniach ewolucji systemów zapylania
Barszcz zwyczajny, podobnie jak inne gatunki z rodziny baldaszkowatych, jest uważany za prymitywny pod względem zapylania – jego kwiaty są odwiedzane przez wiele gatunków owadów, które wielu autorów utożsamia z zapylaczami (Corbet 1970; Sheppard 1991; Proctor i in. 1996). Pogląd ten jest często kwestionowany (Grace, Nelson 1981; Zych 2002, 2004). Ze względu na duże zróżnicowanie morfologiczne, H. sphondylium jest doskonałym modelem do badań nad ewolucją systemów zapylania. W zależności od ujęcia taksonomicznego, dzieli się go na kilka-, kilkadziesiąt podgatunków, odmian i form, z których większość różni się między sobą budową kwiatów i kwiatostanów. Niektóre z tych taksonów są czasem traktowane jako odrębne gatunki. Spośród nich w Polsce występują dwa podgatunki: nominatywny (H. sphondylium subsp. sphondylium) i syberyjski (H. sphondylium subsp. sibiricum). Pierwszy z nich ma białe kwiaty, z których większość charakteryzuje się symetrią dwuboczną. Baldachy podgatunku nominatywnego są gęste i tworzą jedną powierzchnię. Kwiaty podgatunku syberyjskiego są żółtozielone i promieniste, zaś jego kwiatostany mają luźną strukturę (Gawłowska 1961; Sheppard 1991; Zych 2002). Jednym z niewielu stanowisk w Polsce, na których występują obydwa taktony, jest Wigierski Park Narodowy. Podobne różnice w budowie kwiatów i kwiatostanów występują pomiędzy innymi gatunkami lub podgatunkami europejskich barszczy. W badaniach nad biologią zapylania takie zróżnicowanie cech interpretuje się zwykle jako adaptację do przywabiania różnych grup zapylaczy. Zjawisko to jest postrzegane jako istotny czynnik ewolucji okrytozalążkowych (Kay 1976; Waser, Price 1983; Schemske, Bradshaw 1999; Bradshaw, Schemske 2003; patrz także: Herrera 1996). Uważa się, że białe kwiaty są bardziej atrakcyjne dla muchówek, zaś zielonkawe dla chrząszczy (Faegri, van der Pijl 1979; Willemstein 1987; Proctor i in. 1996).
Owady odwiedzające kwiaty
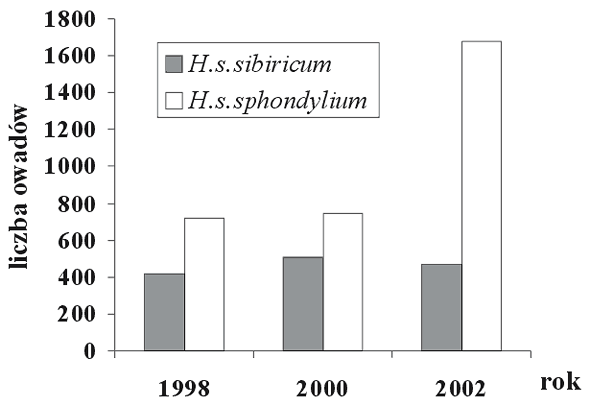
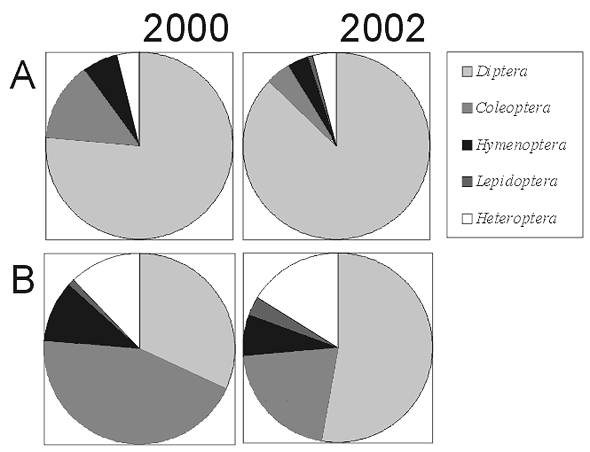
Baldachy barszczu zwyczajnego są atrakcyjne dla wielu grup owadów odwiedzających je w poszukiwaniu pyłku i nektaru (Zych 2002, 2003a). Entomofauna kwiatostanów H. sphondylium, badana na dwóch stanowiskach w Wigierskim Parku Narodowym (tab. 1), obejmuje ponad 107 gatunków należących do siedmiu rzędów; główną grupę stanowią muchówki i chrząszcze (Zych 2003a; tab. 2). Jak wynika z porównania liczby wizyt, białe kwiatostany podgatunku nominatywnego są znacznie atrakcyjniejsze dla owadów niż zielonkawe baldachy barszczu syberyjskiego (rys. 1). Zwraca także uwagę większy procentowy udział chrząszczy na zielonkawych kwiatach barszczu syberyjskiego (rys. 2), co potwierdzałoby wspomniane wcześniej przypuszczenia literaturowe.
Rys. 1. Liczba owadów odwiedzających kwiatostany dwóch podgatunków barszczu zwyczajnego, H. sphondylium subsp. sphondylium i H. sphondylium subsp. sibiricum, na terenie Wigierskiego Parku Narodowego w trakcie trzech sezonów badawczych. We wszystkich sezonach obserwacje dla każdego podgatunku prowadzono w czterech dwunastogodzinnych sekwencjach (szczegóły zob. Zych 2002)
Rys. 2. Procentowy udział owadów z różnych rzędów w odwiedzinach na kwiatostanach w fazie żeńskiej dwóch podgatunków H. sphondylium na terenie Wigierskiego Parku Narodowego w sezonach badawczych 2000 i 2002 (A – H. sphondylium subsp. sphondylium, B – H. sphondylium subsp. sibiricum)
Zapylacze
Odwiedzanie kwiatów danej rośliny przez dużą liczbę gatunków owadów nie oznacza jednak, że wszystkie z nich są równie cenne jako zapylacze (Waser i in. 1996; Pellmyr 2002). W przypadku opisywanego tutaj H. sphondylium grupę tę stanowi zaledwie kilka gatunków, głównie muchówek (Eriozona syrphoides, Lucilia spp., Thricops nigrifrons) i chrząszczy (Stenurella spp.) – w dodatku grupy zapylaczy są prawie jednakowe dla obu występujących na terenie WPN podgatunków (Zych 2002, 2003a). Oznacza to, że występowanie barwnych morf jest już tylko prawdopodobnie pozostałością historyczną i wynika z działania dawnych nacisków selekcyjnych, które istniały w miejscu powstania badanych taksonów. Podobnie jest w przypadku kontrastującego, ciemnofioletowego kwiatu, wytwarzanego w środku kwiatostanów marchwi zwyczajnej (Daucus carota), należącej do tej samej rodziny baldaszkowatych; próby wyjaśnienia jego funkcji nie powiodły się (Lamborn, Ollerton 2000). Mimo to interesujące jest prześledzenie ewolucji taksonów Heracleum z uwzględnieniem cech, które są odpowiedzialne za przywabianie zapylaczy, ale także tych, stanowiących ważne cechy diagnostyczne w oznaczaniu badanej grupy.
Filogeneza
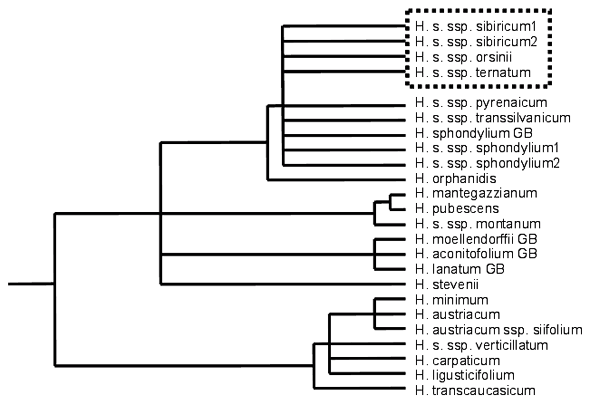
W przypadku prób rekonstrukcji ewolucji systemów reprodukcyjnych blisko spokrewnionych taksonów, główną przeszkodą jest zwykle ustalenie stosunków pokrewieństwa między nimi (Weller, Sakai 1999 i lit. cyt.), szczególnie wtedy, kiedy obiekty uznawane są za „taksonomicznie trudne”. Do takich można zaliczyć rodzaj Heracleum. Podobnie jak w przypadku innych baldaszkowatych (Heywood 1971), jego systematyka jest dość zawiła i do niedawna brak było nowych opracowań dotyczących filogenezy tego rodzaju. Dopiero zastosowanie nowoczesnych metod, opartych na analizie markerów molekularnych (w tym przypadku jądrowych sekwencji ITS rDNA), pozwoliło uzyskać dość miarodajne wyniki (Zych 2003b; rys. 3). Badania wykazały, że grupę blisko spokrewnioną stanowią europejskie barszcze – endemity różnych masywów górskich. Wszystkie, z wyjątkiem krymskiego barszczu Stevena (H. stevenii) oraz macedońskiego H. orphanidis, lokują się na tej samej gałęzi. Pozycje tę potwierdzają również badania biometryczne. Niestety odcinki ITS większości podgatunków H. sphondylium charakteryzowały się identyczną sekwencją nukleotydów. Uniemożliwiło to ustalenie wzajemnych relacji pokrewieństwa w tej gałęzi. Podobnie jak podgatunki H. sphondylium – identyczne sekwencje ITS miały także: barszcz omszony (H. pubescens) – endemit Krymu – oraz barszcz kaukaski (H. mantegazzianum; sekwencja tego gatunku została ustalona na podstawie materiałów zielnikowych zebranych w Wigierskim Parku Narodowym) – gatunek silnie inwazyjny w całej Europie. Niewykluczone, na co wskazują także badania morfologiczne, że stanowią one ten sam gatunek (Zych 2003a, 2003b). Aby prześledzić ewolucję poszczególnych cech, należało dokonać ich mapowania na drzewach filogenetycznych. Przeprowadzone analizy wykazały, że kształt blaszki liściowej, uznawany za ważny znacznik taksonomiczny i używany jako podstawa do konstrukcji kluczy do oznaczania gatunków (Mandenova 1951, 1962; Brummitt 1968), jest cechą silnie homoplastyczną. W świetle analiz poszczególne stany tej cechy – liść niepodzielony, pierzastosieczny, trójlistkowy czy nieparzystopierzasty – nie wyznaczają grup wywodzących się od jednego przodka (Zych 2003a).
Rys. 3. Uproszczone drzewo filogenetyczne europejskich przedstawicieli rodzaju Heracleum L., uzyskane na podstawie jądrowych sekwencji ITS rDNA (za Zych 2003a) (przerywaną linią obwiedzione są taksony o promienistych żółtozielonych kwiatach, pozostałe gatunki mają kwiaty białe o symetrii dwubocznej)
Z powodu braku rozdziału w gałęzi Heracleum sphondylium trudno niestety jednoznacznie wnioskować o ewolucji cech kwiatów. Można przypuszczać, że żółtozielona barwa, charakterystyczna m.in. dla podgatunku syberyjskiego, jest cechą pochodną ewolucyjnie – poza kilkoma taksonami z grupy Heracleum sphondylium pozostałe barszcze charakteryzują się kwiatami białymi o symetrii dwubocznej (Zych 2003a, rys. 3). Jak zatem mogła wyglądać ewolucja systemów zapylania europejskich barszczy?
Krótka historia barszczy
Badania wskazują, że udział muchówek w zapylaniu roślin wzrasta wraz z wysokością względną i spadkiem temperatury oraz że owady te pełnią ważną rolę w zapylaniu roślin flory arktycznej i górskiej (Kevan 1972; Arroyo i in. 1982). Jak wynika z badań zasięgów badanych podgatunków barszczu zwyczajnego w Polsce, występowanie podgatunku nominatywnego było ograniczone do południowych, górzystych regionów Polski, zaś podgatunek syberyjski występował w części centralnej i północnej (Gawłowska 1956, 1957). Obecnie zasięgi te ulegają wymieszaniu, a oba taksony można znaleźć w różnych regionach kraju (Zych 2002). Na podstawie danych o europejskich zasięgach gatunków Heracleum można stwierdzić, że rodzaj ten skupia głównie gatunki górskie. Analizy filogenetyczne wskazują, że taką strategią życiową charakteryzowały się także taksony ancestralne. W przypadku barszczy europejskich, poza podgatunkiem nominatywnym H. sphondylium, który występuje także na nizinach, wszystkie pozostałe gatunki Heracleum oraz podgatunki H. sphondylium występujące w górach, charakteryzują się białymi kwiatami o symetrii dwubocznej. Analizy filogenetyczne pokazują, że podgatunek syberyjski oraz inne podgatunki z zielonkawożółtymi, promienistymi kwiatami, są formami pochodnymi. Żaden z nich nie jest formą górską. Można przypuszczać, iż w momencie różnicowania się tych taksonów i poszerzania się ich zasięgów, w miejscach ich występowania istniały ważne z punktu widzenia sukcesu reprodukcyjnego grupy owadów, które faworyzowały pewne cechy morfologiczne kwiatów. Wśród roślin górskich, rosnących głównie na otwartej przestrzeni, istnieje duża konkurencja o owady zapylające (Kevan 1972), więc także w przypadku górskich barszczy istniał silny dobór w kierunku utrzymania atrakcyjnych kwiatostanów. Formy nizinne, takie jak subsp. sibiricum, zdobywały nowe siedliska, głównie leśne i zaroślowe, gdzie istniały odmienne grupy zapylaczy, wśród których dużą część mogły stanowić chrząszcze preferujące zielonkawe kwiaty. Pojawiające się spontanicznie rośliny, charakteryzujące się takimi właśnie cechami, odnosiły większy sukces reprodukcyjny, w związku z czym, stawały się w danej populacji coraz częstsze. Nie bez znaczenia mogła być także luźniejsza struktura kwiatostanów, form o zielonożółtych kwiatach. Szczególnie w przypadku chrząszczy, które są mniej aktywne na kwiatach niż na przykład muchówki, mogło to stymulować ruch pomiędzy kwiatostanami blisko rosnących osobników, a wraz z tym zwiększać stopień obcopylności. Takie naciski selekcyjne mogły doprowadzić do powstania istniejących dzisiaj form, a rozszerzanie się zasięgu geograficznego gatunku mogło osłabić działanie doboru kierunkowego i uniemożliwić powstanie bardziej wyspecjalizowanych rozwiązań. Obecna przewaga, jeżeli chodzi o występowanie białokwiatowej formy ancestralnej, wynika z jej większej konkurencyjności o zapylaczy, zwłaszcza w sytuacjach, w których obserwuje się znaczny spadek powierzchni siedlisk leśnych na korzyść obszarów otwartych. Brak bardziej wyspecjalizowanych form związanych z przystosowaniem się do węższej grupy zapylaczy, może wynikać z zalet generalizacji. Strategia taka jest bardziej opłacalna z punktu widzenia bezpieczeństwa reprodukcji. Możliwe, że to także przykład ewolucyjnego „ograniczenia” charakteryzującego niektóre rodziny okrytozalążkowych (Ollerton 1996; Johnson, Steiner 2000), co dla baldaszkowatych postulował już Bell (1971). Ograniczenia takie uniemożliwiają znaczne zmiany ewolucyjne w obrębie kwiatów i kwiatostanów, i pozwalają zaledwie na subtelne zmiany morfologii, obserwowane na przykład wśród amerykańskich gatunków z tej rodziny (Lindsey, Bell 1985). Taki ewolucyjny konserwatyzm jest pomocny w zdobywaniu nowych obszarów, ponieważ w momencie utraty ważnego zapylacza, stratę tę zrekompensować mogą inne grupy owadów odwiedzające kwiaty danej rośliny.
Podsumowanie
Występujące na terenie Wigierskiego Parku Narodowego dwa podgatunki barszczu zwyczajnego: H. sphondylium subsp. sphondylium i H. sphondylium subsp. sibiricum – różnią się budową oraz kolorem kwiatów i kwiatostanów. Pierwszy z nich ma kwiaty białe o symetrii dwubocznej, ułożone w zwarte kwiatostany, kwiaty drugiego są żółtozielone, zebrane w luźne baldachy. Takie zróżnicowanie cech interpretuje się jako przystosowanie do różnych grup zapylaczy. Trzyletnie badania terenowe wykazały, że kwiaty subsp. sphondylium są chętnie odwiedzane przez różne grupy owadów, a baldachy subsp. sibiricum są preferowane przez chrząszcze. Mimo obserwowanych różnic w atrakcyjności, oba taksony mają identyczny zespół zapylaczy (grupa złożona głównie z muchówek Eriozona syrphoides, Lucilia spp.). Badania filogenetyczne, oparte na jądrowych sekwencjach ITS rDNA, wskazują, że barszcze o kwiatach żółtozielonych są grupą bardziej zaawansowaną ewolucyjnie. Brak przełożenia tych wyników na zróżnicowanie systemów zapylania sugeruje, że istnienie barwnych morf jest przystosowaniem do warunków historycznych, panujących w miejscu różnicowania się tych taksonów.
Tabela 1. Opis stanowisk na terenie Wigierskiego Parku Narodowego, gdzie przeprowadzono badania zapylaczy dwóch podgatunków H. sphondylium
Tabela 2. Owady odwiedzające kwiaty Heracleum sphondylium subsp. sphondylium i H. sphondylium subsp. sibiricum na dwóch stanowiskach w Wigierskim Parku Narodowym (tabela zawiera taksony obserwowane i poławiane na roślinach w sezonach 1998, 2000 i 2002; niektóre owady oznaczane były wyłącznie do rangi rodziny)
Diptera – Anthomyidae; Asilidae: Andrenosoma atra (L.); Bombyliidae: Villa hottentotta (L.); Calliphoridae: Calliphora vomitoria (L.), Lucilia caesar (L.), Lucilia illustris (Meig.); Chloropodidae; Conopidae: Conops quadrifasciatus (Deg.); Culicidae; Empididae: Euempis tesselata Fabr.; Fanniidae: Fannia armata (Meig.), inne Fannia (R.-D.); Muscidae: Graphomya maculata (Scop.), Morellia aenescens (R.-D.), Phaonia angelicae (Scop.), Phaonia (R.-D.), Thricops nigrifrons (R.-D.); Sarcophagidae: Sarcophaga carnaria (L.); Sepsidae; Stratiomyidae: formosa (Scop.), Odontomyia hydroleon (L.); Syrphidae: Cheilosia honesta (Rodani), Cheilosia melanopa (Zett.), Cheilosia scutellata (Fall.), Cheilosia pilifera Beck, other Cheilosia Meig., Episyrphus balteatus (De Geer), Episyrphus cinctellus (Zett.), Eriozona syrphoides (Fall.), Eristalis abrustorum (L.), Eristalis horticola (De Geer), Eristalis pertinax (Scop.), Helophilus pendulus (L.), Ischyrosyrphus glaucius (L.), Melanostoma mellinum (L.), Metasyrphus corollae (Fabr.), Metasyprhus lapponicus (Zett.), Myatropa florea (L.), Parasyrphus lineola (Zett.), Parasyrphus vittiger (Zett.), Sphaerophoria menthrasti (L.), Sphaerophoria scripta (L.), Syritta pipiens (L.), Syrphus ribesii (L.), Volucella pelucens (L.), inne Syrphidae; Tabanidae: Tabanus bovinus (L.); Tachinidae: Actia lamma Meig., Ernestia rudis (Fall.), Eurithia anthophila (R.-D.), Eurithia consobrina (Meig.), Hyalurgus lucidus Meig., Phasia hemiptera (Fabr.); inne Diptera
Coleoptera – Cantharidae: Rhagonycha fulva (Scop.); Cerambycidae: Dinoptera collaris (L.), Leptura maculata (Poda), Stenurella bifasciata (O. F. Müller), Stenurella melanura (Vill.); Coccinellidae: Coccinella septempunctata (L.); Melyridae: Dasytes (Paykull); Mordellidae: Mordellistena (Costa); Nitidulidae: Cychramus luteus (Fabr.), Meligethes (Steph.); Oedemeridae: Chrysanthia viridissima (L.); Scarabaeidae: Cetonia aurata (L.)
Hymenoptera – Andrenidae: Andrena minutula (Kirby), Andrena rosae Panz.; Apidae: Bombus lapidarius (L.), Bombus terrestris (L.); inne Apoidea; Argidae: Arge ochropus (Gmelin), Arge ustulata (L.); Cimbicidae: Cimbex Olivier; Colletidae: Hylaeus communis (Nylander), Hylaeus confusus (Nylander); Eumenidae: Eumenes coronatus (Panz.), Eumenes pedunculatus (Panzer), Symmorphus mutinense (Baldini); Halictidae: Lasioglossum calceatum (Scop.); Ichneumonidae; Mellitidae: Macropis fulvipes (L.); Phytophaga – inne; Sphecidae: Crabro cribarius (L.), Crossocerus assimilis (Smith), Crossocerus wesmaeli (V. d. Lind.), Ectemnius lapidarius (Panz.); Vespidae: Dolichovespula sylvestris (Scop.), Vespa crabro (L.); inne Vespoidea;
Lepidoptera – Geometridae; Nymphalidae: Araschnia levana (L.), Argynnis paphia (L.), Erebia medusa (Denis & Schiffermüller); Papilionidae: Papilio machaon (L.); Pieridae: Gonepteryx rhamni (L.), Pieris brassicae (L.); Pterophoridae; Zygaenidae: Zygaena filipendulae (L.), Zygaena trifolii (Esper)
Heteroptera – Miridae: Calocoris affinis (Herrich-Schaff.), Miris striatus (L.), Orthops calmi (L.); Pentatomidae: Palomena viridissima (Poda), inne Pentatomidae
Mecoptera – Panorpidae: Panorpa communis (L.)
Neuroptera – Chrysopidae: Chrysopa pallens (Rambur)
Literatura
Arroyo M. T. K., Primack R., Armesto J., 1982: Community studies in pollination ecology in the high Andes of Central Chile. 1: Pollination mechanisms and altitudinal variation. American Journal of Botany, 69:82–97. Bell C.R., 1971: Breeding systems and floral biology of the Umbelliferae or evidence for specialization in unspecialized flowers. W: Heywood V. H. (red.): The Biology and Chemistry of the Umbelliferae. London, Academic Press, s. 93–107. Bradshaw jr. H. D., Schemske D. W., 2003: Allele substitution at a flower colour locus produces a pollinator shift in monkeyflowers. Nature, 426:176–178. Brummitt R. K., 1968: Heracleum L. W: Tutin T. G. i in. (red.): Flora Europaea, vol. 2. Cambridge, Cambridge University Press, s. 364–366. Corbet S. A., 1970: Insects on hogweed flowers: a suggestion for a student project. Journal of Biological Education, 4:133–143. Faegri K., Pijl L. van der, 1979: The Principles of Pollination Ecology. Oxford, Pergamon Press. Gawłowska M., 1956: Heracleum sphondylium L. i Heracleum sibiricum L. na ziemiach polskich. Dissertationes Pharmaceuticae, 7:141–164. Gawłowska M., 1957: Badania nad rozmieszczeniem Heracleum sphondylium L. i H. sibiricum L. w Polsce. Fragmenta Floristica et Geobotanica, 3:61–68. Gawłowska M., 1961: Systematyka wewnątrzgatunkowa Heracleum sphondylium L. i Heracleum sibiricum L. występujących w Polsce i krajach sąsiednich. Fragmenta Floristica et Geobotanica, 7:1–39. Grace J., Nelson M., 1981: Insects and their pollen loads at a hybrid Heracleum site. New Phytologist, 87:413–423. Herrera C. M., 1996: Floral traits and plant adaptation to insect pollinators: a devil’s advocate approach. W: Lloyd D. G., Barrett S. C. (red.): Floral Biology. Studies on Floral Evolution in Animal-Pollinated Plants. New York, Chapman, Hall, s. 65–87. Heywood V. H., 1971: Systematic survey of Old World Umbelliferae. W: Heywood V. H. (red.): The Biology and Chemistry of the Umbelliferae. London, Academic Press, s. 31–41. Johnson S. D., Steiner K. E., 2000: Generalization versus specialization in plant pollination systems. Trends in Ecology and Evolution, 15:140–143. Kay Q. O. N., 1976: Preferential pollination of yellow-flowered morphs of Raphanus raphanistrum by Pieris and Eristalis spp. Nature, 261:230–232. Kevan P. G., 1972: Insect pollination of high arctic flowers. Journal of Ecology, 60:831–847. Lamborn E., Ollerton J., 2000: Experimental assessment of the functional morphology of inflorescence of Daucus carota (Apiaceae): testing the ‘fly catcher effect’. Functional Ecology, 14:445–454. Lindsey A. H., Bell C. R., 1985: Reproductive biology of Apiaceae. 2: Cryptic specialization and floral evolution in Thaspium and Zizia. American Journal of Botany, 72:231–247. Mandenova I. P., 1951: Heracleum L. W: Shishkin B. K. (red.): Flora SSSR. T. 17. Moskwa–Leningrad, Izdatelstvo Akademii Nauk SSSR, s. 223–259. Mandenova I. P., 1962: Taxonomic review of Turkish species of Heracleum. Notes from the Royal Botanic Garden, 24:173–181. Ollerton J., 1996: Reconciling ecological processes with phylogenetic patterns: the apparent paradox of plant-pollinator systems. Journal of Ecology, 84:767–769. Pellmyr O., 2002: Pollination by animals. W: Herrera C. M., Pellmyr O. (red.): Plant-Animal Interaction. An Evolutionary Approach. Oxford, Blackwell, s. 157–184. Proctor M., Yeo P., Lack A., 1996: Natural History of Pollination. London, Harper Collins Publishers. Schemske D. W., Bradshaw jr. H. D., 1999: Pollinator preference and the evolution of floral traits in monkeyflowers (Mimulus). Proceedings of the National Academy of Sciences, 96:11910–11915. Sheppard A. W., 1991: Heracleum sphondylium L. Journal of Ecology, 79:235–258. Waser N. M., Price M. V., 1983: Pollination behavior and natural selection for flower colour in Delphinium nelsonii. Nature, 302:422–424. Waser N. M., Chittka A. L., Price M. V., Williams N. M., Ollerton J., 1996: Generalization in pollination systems, and why it matters. Ecology, 77:1043–1060. Weller S. G., Sakai A. K., 1999: Using phylogenetic approaches for the analysis of plant breeding system evolution. Annual Review of Ecology and Systematics, 30:167–199. Willemstein S. C., 1987: An evolutionary basis for pollination ecology. Leiden Botanical Series, vol. 10. Leiden, University Press. Zych M., 2002: Pollination biology of Heracleum sphondylium L. (Apiaceae). The advantages of being white and compact. Acta Societatis Botanicorum Poloniae, 71:163–170. Zych M., 2003a: Biologia zapylania i pozycja filogenetyczna dwóch podgatunków barszczu zwyczajnego (Heracleum sphondylium L.). Rozprawa doktorska. Wydział Biologii Uniwersytetu Warszawskiego. Zych M., 2003b: Phylogeny of the European Heracleum L. species as inferred from ITS rDNA sequences. Scripta Botanica Belgica, 27:64. Zych M., 2004: Biologia zapylania baldaszkowatych (Apiaceae) – stare mity i nowe perspektywy. Wiadomości Botaniczne, 48:7–15
|