|
AUGUSTOWSKO-SUWALSKIE TOWARZYSTWO NAUKOWE
Proszę chwilę zaczekać, ładuję stronę... |
|
AUGUSTOWSKO-SUWALSKIE TOWARZYSTWO NAUKOWE
Proszę chwilę zaczekać, ładuję stronę... |
|
Piotr Zieliński, Andrzej Górniak, Joanna Maksimiuk
Letni bakterioplankton jeziora Wigry
Obecnie powszechnie uważa się, że bakterie są niezwykle ważnym ogniwem troficznym w wodach naturalnych, a od procesów mikrobiologicznych zależy prawidłowe funkcjo- nowanie i stabilność ekologiczna wielu ekosystemów wodnych (Chróst 1994). Bakterie pełnią kluczową i dominującą rolę w procesach krążenia materii mineralnej i organicznej w każdym środowisku wodnym (Rheinbeimer 1987; Chróst 1995). Dotychczasowe badania dotyczące bakterioplanktonu jeziora Wigry miały głównie charakter sanitarny (Niewolak 1999). Jezioro Wigry dzięki swojej budowie morfologicznej jest ekosystemem o różnorodnych warunkach dla rozwoju bakterioplanktonu. Ponadto działania ochronne Wigierskiego Parku Narodowego i ich efekty w szybkim tempie wpływają na przywracanie stanu równowagi w jeziorze. Rozczłonkowanie tego jeziora i duże różnice w głębokościach plos, różnice w warunkach wymiany wody, poprawa jakości wody, skłaniają do poznania wpływu tych czynników na bakterioplankton. Jednym z głównych celów pracy była analiza przestrzennego zróżnicowania liczebności bakterioplanktonu jeziora oraz zależności z różnymi formami węgla organicznego i wybranymi parametrami jakości wody.
Rys. 1. Rozmieszczenie stanowisk poboru prób
Teren i metody
Badania mikrobiologiczne i hydrochemiczne wody jeziora Wigry przeprowadzono w drugiej połowie sierpnia 2002 roku podczas długoterminowej suszy atmosferycznej i hydrologicznej. Wodę do analiz pobierano z 15 stanowisk (rys. 1). Próby na każdym stanowisku, za wyjątkiem stanowiska 4b, pobierano z trzech warstw termicznych. Ogólną liczbę bakterii oznaczano metodą mikroskopową z wykorzystaniem roztworu DAPI (Hobbie i in. 1977). Biomasę bakterii obliczano, wykorzystując metodę zaproponowaną przez Theil-Nielsena i Sondergaarda (1998). Analizy chemiczne wody wykonano zgodnie metodami opisanymi przez Hermanowicza i in. (1976) oraz Górniaka (w tym tomie). Rozpuszczony węgiel organiczny oznaczano metodą katalitycznego spalania w wysokiej temperaturze za pomocą analizatora węgla firmy Shimadzu TOC-5050A. Jakość rozpuszczonej materii organicznej opisano za pomocą wskaźnika aromatyczności węgla organicznego SUVA (Rostan i Cellot 1995, zm.).
Wyniki
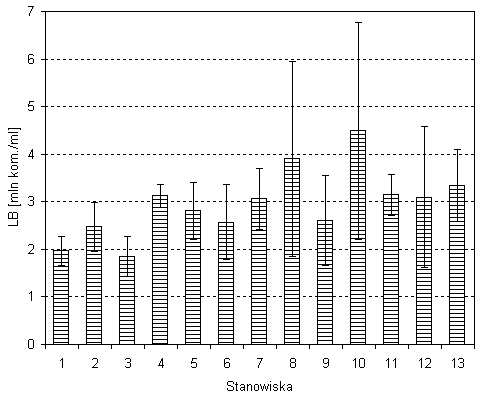
W sierpniu 2002 roku średnia liczebność bakterii (LB) w jeziorze Wigry wynosiła 3,1 mln kom./cm3. Średnia liczebność bakterii na poszczególnych stanowiskach była podobna, pomijając stanowiska znajdujące się blisko brzegów (stanowisko 8, 10). Średnia wartość LB wzrastała od plosa północnego w kierunku plosa zachodniego (za wyjątkiem stanowiska 4b w Zatoce Hańczańskiej, znajdującego się tuż przy ujściu do jeziora Czarnej Hańczy). Maksymalne zagęszczenie odnotowano na stanowisku nr 10 w rejonie wyspy Ostrów (rys. 2). Minimalna liczebność występowała na stanowisku nr 3 w Plosie Wigierskim (rys. 2). Istotnie statystycznie różnice wartości LB odnotowano między stanowiskami 1 i 10 oraz 3 i 10.
Rys. 2. Średnia liczebność bakterii na poszczególnych stanowiskach jeziora Wigry
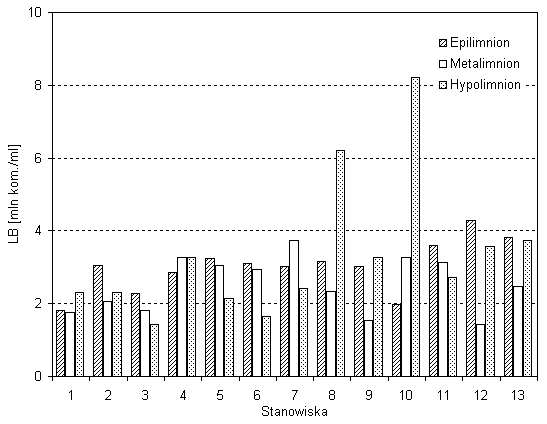
Zagęszczenia bakterii w epilimnionie (3,17 mln kom./cm3) i hypolimnionie (3,21 mln kom./cm3) jeziora było zbliżone, natomiast w metalimnionie było mniejsze o ok. 0,5 mln kom./cm3. Liczebność bakterii w epilimnionie jeziora była najmniej zmienna (CV=25proc.) na badanych stanowiskach (rys. 3). Maksymalną LB w epilimnionie odnotowano na stanowisku nr 12 w zatoce Wigierki, a minimalną na stanowisku nr 1 w zatoce Zadworze (rys. 3). W metalimnionie liczebność bakterii była bardziej zmienna na poszczególnych stanowiskach (CV=34 proc.) (rys. 3). Największe zagęszczenie komórek bakterii występowało w plosie środkowym na stanowisku 7 (3,73 mln kom./cm3), a najmniejsze w zatoce Wigierki na stanowisku nr 12 (1,42 mln kom./cm3) (rys. 3).
Rys. 3. Średnia liczebność bakterii w różnych warstwach
termicznych na poszczególnych stanowiskach
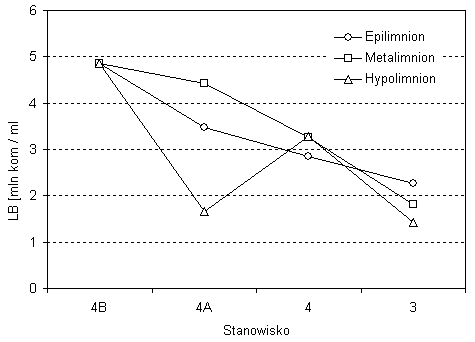
Hypolimnion jeziora Wigry charakteryzował się największą liczebnością bakterioplanktonu (3,21 mln kom./cm3) i największą jego zmiennością (CV=58,5proc.) W jeziorze Wigry można wyróżnić dwa typy hypolimnionu: płytki i głęboki. W płytkim hypolimnionie, do 30 m, maksymalną liczebność bakterii odnotowano na stanowisku nr 10 w rejonie wyspy Ostrów wynoszącą aż 8,21 mln kom./cm3. Minimalną natomiast wartość LB stwierdzono w Zatoce Hańczańskiej na stanowisku nr 4a (1,66 mln kom./cm3). W głębokim hypolimnionie o głębokości powyżej 30 m największą koncentrację komórek bakteryjnych w wodzie odnotowano na stanowisku nr 8 w Zatoce Bielańskiej (6,21 mln kom./cm3), a najniższą, wynoszącą 1,44 mln kom./cm3, w Plosie Wigierskim (stanowisko nr 3) (rys. 3). W jeziorze Wigry widoczne były cztery typy pionowego rozmieszczenia bakterioplanktonu. W części północnej Wigier ilość bakterii przeważnie zmniejszała się wraz z głębokością (stanowisko 3, 5, 6 i 11). Tylko w dwóch przypadkach sytuacja była odwrotna, na stanowiskach nr 4 i 10, gdzie zagęszczenie bakterii było największe w hypolimnionie i zmniejszało się ku powierzchni zbiornika. Na pozostałych badanych stanowiskach maksymalną lub minimalną liczebność bakterii odnotowano w metalimnionie (rys. 3). W przepływowej części Wigier, tj. w Zatoce Hańczańskiej, średnia liczebność bakterii zmniejszała się wzdłuż przepływu Czarnej Hańczy od ujścia do plosa północnego (rys. 4). Sytuacja taka utrzymywała się głównie w epilimnionie i metalimnionie tej części jeziora Wigry (rys. 4). W hypolimnionie liczebność bakterii początkowo spadała z 4,8 mln kom./cm3 do 1,66 mln kom./cm3, następnie wzrastała i ponownie obniżyła się do minimum na stanowisku nr 3.
Rys. 4. Liczebność bakterii w warstwach termicznych
wody wzdłuż przepływu Czarnej Hańczy
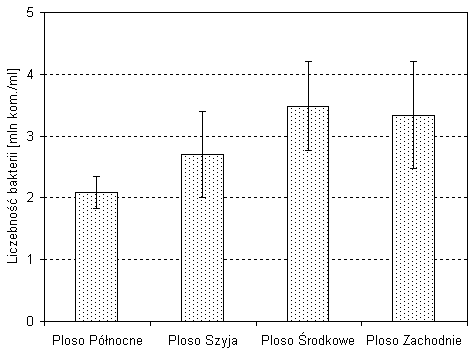
Istnieje istotne zróżnicowanie liczebności bakterii w poszczególnych częściach Wigier. Największe zagęszczenie bakterii występowało w plosie środkowym i zachodnim, natomiast najmniejsze w plosie północnym i Szyja (rys. 5). W plosie zachodnim stwierdzono maksymalną koncentrację bakterii w epilimnionie, a w plosie środkowym maksymalną ilość bakterii w hypolimnionie. Istotne statystycznie różnice odnotowano pomiędzy LB w plosie północnym i środkowym, a także pomiędzy LB w plosie północnym i zachodnim (rys. 5).
Rys. 5. Średnia liczebność bakterii w różnych częściach jeziora Wigry
W sierpniu 2002 roku średnia biomasa bakterii wynosiła 0,11 mgC/dm3, co stanowiło 5 proc. średniego stężenia cząsteczkowego węgla organicznego (POC) w wodach jeziora. Największą część POC stanowiła biomasa bakterii na stanowisku nr 12 w zatoce Wigierki, natomiast najmniejszą w zatoce Zadworze (tab. 1). W plosie środkowym stwierdzono największy udział biomasy bakterioplanktonu w tworzeniu POC, wynoszący prawie 20 proc. W plosie północnym udział ten był najmniejszy, równy 3,55 proc. (tab. 1).
Tabela 1. Średnie stężenie POC, biomasa bakterii (BB) oraz udział bakterii w tworzeniu POC w poszczególnych plosach jeziora Wigry w sierpniu 2002 roku
Pod względem hydrochemicznym jezioro Wigry wykazywało charakter mezotroficzny ewentualnie lekko eutroficzny (w części północnej) (tab. 2). Poszczególne plosa nie wykazały istotnego zróżnicowania pod względem zawartości TP, jonów NO3- i parametru SWWT. Znaczne różnice wykazywały zawartości: rozpuszczonego węgla organicznego (DOC), żelaza, jonów NH4 i SO42-, a także stopnia wysyceniem wody tlenem (WT) i wskaźnika aromatyczności rozpuszczonego węgla organicznego (SUVA).
Tabela 2. Średnie wartości (z całego słupa wody) parametrów fizykochemicznych wody na poszczególnych stanowiskach jeziora Wigry (sierpień 2002). Przew. – przewodność właściwa wody, WT – wysycenie wody tlenem
Dyskusja
Liczebność bakterii i innych drobnoustrojów w wodzie na ogół wzrasta wraz ze wzrostem trofii jeziora (Chróst 1995). W jeziorach oligotroficznych ogólna liczba bakterii w 1 cm3 wynosi blisko 500 tysięcy komórek, w mezotroficznych waha się od 500 tysięcy do 1,5 mln, w eutroficznych osiąga średnio 2–4 mln kom., a w niektórych przewyższa nawet 10 mln (Kuźniecow 1970, za Wetzel 2001). Jezioro Wigry charakteryzuje się wysoką liczebnością bakterioplanktonu w porównaniu z innymi jeziorami mezotroficznymi regionu. Jezioro Wigry pod względem liczebności bakterii w wodzie należałoby zaliczyć do jezior eutroficznych, ponieważ ilość bakterii dwukrotnie przekracza 1,5 mln kom./cm3, ale być może w czasie badań odnotowano stan przejściowy, zwłaszcza że jakość wody wskazuje na mezotroficzny charakter wód jeziora (tab. 2). W sierpniu 2002 roku wystąpiły szczególne warunki meteorologiczne (susza hydrologiczna) i obniżony stan wody w jeziorze, które prawdopodobnie przyczyniły się do okresowego wzrostu liczebności bakterioplanktonu w wodzie. Istniały w tym czasie sprzyjające warunki do wzbogacenia wody w łatwo przyswajalne substancje organiczne, gromadzące się w wodzie głównie wiosną i pod koniec lata. Przy wyższej temperaturze powierzchniowej warstwy wody ilość bakterii w wodzie osiąga swoje maksimum (Donderski 1983; Rheinheimer 1987). Porównując średnią liczebność bakterii jezior mezotroficznych w Polsce i na świecie, jezioro Wigry posiada zbliżone wartości LB. Podobne wartości dla zbiorników o niskiej trofii uzyskali: Crisman i in. (1984), Laybourn-Parry i in. (1994), Świątecki (1997), Górniak i Świątecki (1999). Liczebność bakterii w jeziorach mazurskich o różnej trofii była kilkakrotnie większa i wahała się pomiędzy 18,7 mln kom./cm3 a 26,0 mln kom./cm3 (Chróst 1995). Jedynie dane Kuźniecowa (1970, za Wetzel 2001), dotyczące kilku rosyjskich jezior mezotroficznych, nie przekraczały wartości 1,5 mln kom./cm3. Liczebność bakterii w jeziorze Wigry musi być jednak mniejsza niż wskazują wyniki badań prezentowanych w niniejszej pracy. Metoda DAPI zawyża bowiem wartość rzeczywistego zagęszczenia aktywnych fizjologicznie bakterii (Sondergaard, Danielsen 2001). Odmienny charakter północnej od południowo-zachodniej części jeziora Wigry pod względem morfometrii, zagospodarowania zlewni (lasy, pola uprawne, łąki), dopływów i odpływu (Czarna Hańcza) znajduje odzwierciedlenie w różnym stopniu zagęszczenia bakterioplanktonu (rys. 5), co potwierdzają wcześniejsze badania Niewolaka (1998, 1999). W latach 1994–1996 w przedłużeniu Zatoki Hańczańskiej w kierunku półwyspu Rosochaty Róg w wodzie Plosa Wigierskiego oraz w plosie Szyja liczebność letniego bakterioplanktonu zmniejszała się. Według Niewolaka (1999) wody w południowo-zachodniej części jeziora Wigry były czyste lub nieznacznie zanieczyszczone (stanowiska 8–13). Powodowane to było brakiem większych skupisk ludności w zlewni i dopływów do zbiornika. Obecnie największa koncentracja bakterioplanktonu latem 2002 roku w plosie środkowym i zachodnim świadczy o dużej odrębności tej części jeziora, wynikającej głównie z jego specyfiki morfologicznej. W japońskim jeziorze Biwa (zróżnicowanym pod względem budowy misy jeziornej), północna część zbiornika o mezotroficznym charakterze zawierała od 1,6 mln do 3,4 mln/cm3 bakterii, z kolei w mezoeutroficznej, południowej części było ich kilkadziesiąt razy więcej (Nagata 1984). Odnotowany kilka razy większy udział bakterii w tworzeniu POC w południowej części jeziora niż w części północnej (tab. 1) świadczy o warunkach bardziej sprzyjających dla rozwoju bakterioplanktonu w wodach plosa środkowego i zachodniego. Może to być także efekt odmienności fizjologicznej grup bakterii wykorzystujących różny rodzaj materii organicznej. Wyższe zagęszczenie bakterioplanktonu w rejonie Zatoki Hańczańskiej jest wynikiem wnoszonego ładunku nutrientów, a także specyficznego zespołu mikroorganizmów pochodzenia glebowego oraz z oczyszczalni ścieków (rys. 2, 4). Znaczna liczba ekosystemów lotycznych charakteryzuje się większą liczebnością bakterioplanktonu i dużą zmiennością tego parametru w czasie, w porównaniu do jezior (Edwards 1987). Bakterioplankton docierający do jeziora wraz z wodami Czarnej Hańczy ulega stopniowemu rozcieńczeniu, a typowo glebowe bakterie obumierają. W połowie lat 90. maksymalne LB występowały w wodzie Zatoki Hańczańskiej, co należało wiązać z większym niż obecnie dopływem zanieczyszczeń Czarną Hańczą (Niewolak 1999). W rezultacie woda w tej części jeziora Wigry wykazywała nie tylko wyższy stopień eutrofizacji, ale również gorszy stan sanitarno-bakteriologiczny (Zdanowski i in. 1992). Badania te prowadzone były przed modernizacją suwalskiej oczyszczalni ścieków, która po roku 1997 zdecydowanie poprawiła jakość wód Czarnej Hańczy (Zadanowski, Hutorowicz 1997; Zdanowski 1999). Dlatego obecnie notujemy w tej części jeziora mniejsze liczebności bakterii, a jakość wód dopływających uległa znaczącej poprawie w stosunku do lat poprzednich. Zróżnicowanie pionowego rozmieszczenia bakterii (rys. 3) zależy od cech morfologicznych jeziora, objętości i ruchów wody oraz od ilości i jakości dopływów. Pod względem budowy misy jeziornej, jezioro Wigry jest bardzo urozmaicone. Okresowa pionowa cyrkulacja wody, powodująca wymieszanie mas wodnych, zmienia ewentualne różnice w pionowym rozmieszczeniu bakterii (Niewolak 1999). W jeziorach mezotroficznych i eutroficznych, o niewielkiej głębokości, pionowe rozmieszczenie bakterii jest mniej lub bardziej równomierne. Jedynie w okresie zakwitów wody i masowego obumierania fitoplanktonu znaczne zwiększenie liczebności bakterii występuje w warstwie skoku termicznego lub w profundalu, jak miało to miejsce w plosie środkowym i zachodnim (rys. 3). W jeziorze Wigry nie stwierdzono istotnej statystycznie zależności między maksymalną liczebnością bakterii i fitoplanktonem (stężeniem chlorofilu a). Zróżnicowane, pionowe rozmieszczenie bakterii w jeziorze jest związane ze specjalizacją w zakresie wykorzystania różnych źródeł energii, co potwierdzają przeprowadzone analizy statystyczne. W warstwie epilimnionu odnotowano dodatnią korelację LB i stężenia NO3 (r=0,57; p<0,05). Podobne zależności są stwierdzane jedynie w wodach o niskiej trofii (Górniak, Świątecki 1999). W metalimnionie prawdopodobnie wykorzystywane są efektywnie związki fosforu, o czym świadczy dodatnia zależność pomiędzy liczebnością bakterii a stężeniem TP. Fosfor jest czynnikiem przyspieszającym rozkład materii organicznej (Rheinheimer 1987). W metalimnionie jeziora Wigry odnotowano odwrotnie proporcjonalną zależność pomiędzy liczebnością bakterii i wskaźnikiem aromatyczności węgla organicznego (SUVA) (r= –0,65; p<0,05). Rodzaj materii organicznej, a nie jej ilość, jest czynnikiem wpływającym na zgęszczenie bakterioplanktonu. Najatrakcyjniejszą dla bakterii strukturą węgla charakteryzował się DOC ze stanowisk płytszych. W hypolimnionie Wigier (z okresowym deficytem tlenu) zachodzą procesy utleniania siarkowodoru (sulfurykacja), czego dowodem jest dodatnia korelacja pomiędzy LB i siarczanami (r=0,43; p<0,05). Rozpuszczona materia organiczna jest dominującą frakcją związków organicznych w wodzie jeziora Wigry (Górniak, Zieliński 1999). Stanowi ona bazę substratową i energetyczną dla bakterii heterotroficznych, wolnopływających, epifitycznych lub osiadłych na cząstkach detrytusu i zawiesiny nieorganicznej (Chróst 1995). DOC autochtoniczny to w dużej mierze węgiel związków o małej masie cząsteczkowej i nieskomplikowanej strukturze chemicznej (np. cukry proste, aminokwasy i aminy, krótkie peptydy, kilkuwęglowe kwasy organiczne itp.) (Chróst 1994; Chróst, Gajewski 1995; Zieliński, Górniak 1999). W badaniach nie stwierdzono korelacji między zagęszczeniem bakterii a stężeniem rozpuszczonej materii organicznej. Niewielka zależność występuje jedynie pomiędzy LB i DOC w warstwie powierzchniowej (R2=0,22; p<0,05). W jeziorach tego typu większość bakterioplanktonu jest związana z cząsteczkowym węglem organicznym (POC). Szybkie wychwytywanie cząstek o wysokiej jakości powoduje, że materiał ten jest usuwany z wody w stosunkowo małych odległościach od jego źródeł (Chróst 1990, 1993). W badaniach mikrobiologicznych prowadzonych latem 2002 roku nie stwierdzono istotnej statystycznie zależności między liczebnością bakterii a wskaźnikiem trofii Carlsona (TSI), podczas gdy zależności takie stwierdzono np. w jeziorach mazurskich (Chróst 1995). W plosie północnym odnotowano dodatnie korelacje LB i SO4 (r=0,60; p<0,05), a także LB i chlorków (r=0,57; p<0,05). Liczebność bakterii jest proporcjonalna do stopnia zanieczyszczenia wód. Oba parametry są typową miarą antropopresji. W plosie zachodnim dodatnie zależności odnotowano pomiędzy liczebnością bakterii a formami fosforu: PO4 (r=0,71; p<0,05) i TP (r=0,79; p<0,05), a także pomiędzy LB a stężeniem jonów amonowych (r=0,8; p<0,05). Może być to spowodowane intensywnymi przemianami materii organicznej, zachodzącymi w tej części jeziora w warunkach silnego zasilania wodami podziemnymi, bogatymi w tlen (Bajkiewicz-Grabowska 1999). Reakcje redukcji węgla organicznego wymagają także obecności biogenów. Brak bezpośredniej zależności pomiędzy LB a DOC może wynikać ze zbyt szybkich przemian materii organicznej. W kolejnych badaniach należałoby zwrócić uwagę na ewentualną zależność pomiędzy stężeniem DOC a aktywnością bakterioplanktonu. Podsumowując, można stwierdzić, że przestrzenne zróżnicowanie ogólnej liczebności bakterioplanktonu w jeziorze Wigry w większym stopniu nawiązuje do morfologii i hydrologii plos, a w mniejszym stopniu do statusu troficznego wód.
Autorzy dziękują pracownikom pracowni naukowej Wigierskiego Parku Narodowego za pomoc w zebraniu prób.
Abstract
Summer bacterioplankton of Lake Wigry Bacterial abundance (DAPI staining method), dissolved organic carbon (DOC), particulate organic carbon (POC) and water quality were investigated in samples from lake Wigry, NE Poland. Bacterial cell numbers (BCN) in the lake were 3–4 times higher than in other mesotrophic natural lakes. We observed statistically important differences between northern and southern part of lake Wigry. Lower BCN were observed in deeper part of the lake. Some correlations between BCN and water quality parameters were observed (NO3, TP, SO4, Cl) and specific UV absorbance (SUVA). Key factor determining BCN is lake morphology and hydrology more than water chemistry.
Literatura
Bajkiewicz-Grabowska E., 1999: Bilans wodny jeziora Wigry w latach 1981–1995. W: Zdanowski B., Kamiński M., Martyniak A. (red.): Funkcjonowanie i ochrona ekosystemów wodnych na obszarach chronionych. Olsztyn, Wyd. IRŚ, s. 113–128. Chróst R. J., 1990: Microbial ectoenzymes in aquatic environments. W: Overbeck J., Chróst R. J. (red.): Aquatic microbial ecology: biochemical and molecular approaches. New York–Berlin, Springer-Verlag, s. 47–78. Chróst R. J., 1993: Microbial decomposition of organic matter. Importance for functioning of the lake ecosystems. W: Burchardt L. (red.): Interdisciplinary investigations of freshwater ecosystems. Poznań, Sorus, s. 25–38. Chróst R. J., 1994: Microbial enzymatic degradation and utilization of organic matter. W: Overbeck J., Chróst R. J. (red.): Microbial ecology of Lake Plußsee. New York–Berlin, Springer-Verlag, s. 118–174. Chróst R. J., 1995: Znaczenie procesów mikrobiologicznych dla intensywności występowania symptomów eutrofizacji wód. W: Zalewski M. (red.): Procesy biologiczne w ochronie i rekultywacji nizinnych zbiorników zaporowych. Łódź, PIOŚ, s. 71–82. Chróst R. J., Gajewski A., 1995: Microbial utilization of lipids in lake water. FEMS Microbiol. Ecol., 18:45–50. Cirsman T. L., Scheuerman P., Bienert R. W., Beaver J. R., Bays J. S., 1984: A preliminary characterization of bacterioplankton seasonality in subtropical Florida lakes. Verh. Internat. Verein. Limnol., 22:620–626. Donderski W., 1983: Tlenowe bakterie heterotroficzne jezior o różnej trofii. Toruń, UMK. Edwards R. T., 1987: Sestonic bacteria as a food source for filtering invertebrates in two southeastern blackwater rivers. Limnology and Oceanography, 32:221–234. Górniak A., 2004: Zaawansowanie dystrofii sucharów Wigierskiego Parku Narodowego (w tym tomie). Górniak D., Świątecki A., 1999: Mikroorganizmy wodne jeziora Hańcza. W: Zdanowski B., Kamiński M., Martyniak A. (red): Funkcjonowanie i ochrona ekosystemów wodnych na obszarach chronionych. Olsztyn, Wyd. IRŚ, s. 305–315. Górniak A., Zieliński P., 1999: Rozpuszczone związki węgla organicznego w jeziorze Wigry. W: Zdanowski B., Kamiński M., Martyniak A. (red.): Funkcjonowanie i ochrona ekosystemów wodnych na obszarach chronionych. Olsztyn, Wyd. IRŚ, s. 141–152. Hermanowicz W., Dożańska W., Dojlido J., Koziorowski B., 1976: Fizyczno-chemiczne badanie wody i ścieków. Warszawa, Arkady. Hobbie J. E., Daley R. J., Jasper S., 1997: Use of nucleopore filters for counting bacteria by fluorescence microscopy. Appl. Environ. Microbiol., 33:1225–1228. Laybourn-Parry J., Walton J. M., Young R. I., Jones R. I., Shine A., 1994: Protozooplankton and bacterioplankton in a large oligotrophic lake – Loch Ness. Scotland J., Plankton Res., 16:1655–1670. Nagata T., 1984: Bacterioplankton in lake Biwa: Annual fluctuations of bacterial numbers and their possible relationship with environmental variables. Jpn. J. Limnol., 45:126–133. Niewolak S., 1998: The evaluation of the contamination degree and the sanitary and bacteriological state of the waters in the Czarna Hancza river in the region of Suwalki and Wigry National Park. Polish Journal of Environmental Studies, 7:229–241. Niewolak S., 1999: Ocena stopnia zanieczyszczenia i stanu sanitarno-bakteriologicznego wód powierzchniowych i podziemnych na obszarze Wigierskiego Parku Narodowego w latach 1994–1996. W: Zdanowski B., Kamiński M., Martyniak A. (red.): Funkcjonowanie i ochrona ekosystemów wodnych na obszarach chronionych. Olsztyn, Wyd. IRŚ, s. 231–258. Rheinheimer G., 1987: Mikrobiologia wód. Warszawa, PWRiL. Rostan J. C., Cellot B., 1995: On the use of UV spectrophotometry to assess dissolved organic carbon origin variations in the Upper Rhône river. Aquatic Sciences, 57:70–80. Sondegaard M., Danielsen M., 2001: Active bacteria (CTC+) in temperate lakes: temporal and cross-system variations. Journal of Plankton Research, 11:1195–1206. Świątecki A., 1997: Spatial and seasonal changes in bacterioplankton of Konińskie lakes. Arch. Ryb. Pol., 5:173–188. Theil-Nielsen J., Sondergaard M., 1998: Bacterial carbon biomass calculated from biovolumes. Arch. Hydrobiol., 141:195–207. Wetzel R. G., 2001: Limnology. Lake and river ecosystems. New York, Academic Press. Zdanowski B., 1999: Eutrofizacja jezior Wigierskiego Parku Narodowego: zagrożenia i ocena. W: Zdanowski B., Kamiński M., Martyniak A. (red.): Funkcjonowanie i ochrona ekosystemów wodnych na obszarach chronionych. Olsztyn, Wyd. IRŚ, s. 261–277. Zdanowski B, Karpiński A., Prusik S., 1992: Warunki środowiskowe wód jezior Wigierskiego Parku Narodowego. W: Zdanowski B. (red.): Jeziora Wigierskiego Parku Narodowego. Stan eutrofizacji i kierunki ochrony. Zeszyty Nauk. PAN, Człowiek i Środowisko, Wrocław, 3:35–62. Zdanowski B., Hutorowicz A., 1997: Charakterystyka hydrochemiczna Czarnej Hańczy i wpływ tej rzeki na wody jezior Hańcza i Wigry. W: Zintegrowany monitoring środowiska przyrodniczego. Stacja bazowa Wigry (Wigierski Park Narodowy). Warszawa, Biblioteka Monitoringu Środowiska, s. 29–47. Zieliński P., Górniak A., 1999: Analizy rozpuszczonego węgla organicznego w wodach naturalnych. Aparatura Badawcza i Dydaktyczna, 3:37–45.
|