|
AUGUSTOWSKO-SUWALSKIE TOWARZYSTWO NAUKOWE
Proszę chwilę zaczekać, ładuję stronę... |
|
AUGUSTOWSKO-SUWALSKIE TOWARZYSTWO NAUKOWE
Proszę chwilę zaczekać, ładuję stronę... |
|
Anna Kuczyńska, Stanisław Niewolak
Występowanie bakterii heterotroficznych w wodzie jezior dystroficznych na obszarze Wigierskiego Parku Narodowego
Jedną z osobliwości Wigierskiego Parku Narodowego jest względnie duża liczba (20) jezior dystroficznych, w tym kilka objętych ścisłą ochroną (Zdanowski 1999). Wody tych jezior charakteryzują się wysoką koncentracją substancji humusowych, będących produktem częściowego rozkładu roślinności pochodzenia torfowego, głównie Sphagnum, otaczającej zbiorniki tzw. płem, oraz igliwia z okolicznych lasów sosnowo-świerkowych. Wynikiem jest silne zakwaszenie wody (Stangenberg 1936). Wskutek absorpcyjnych właściwości kwasów humusowych, wody te są ubogie w N i P oraz sole mineralne. Brunatne zabarwienie wody tych zbiorników ogranicza penetrację promieni świetlnych i produkcję pierwotną. Częstym zjawiskiem w tych jeziorach są deficyty tlenowe i obecność siarkowodoru. Wszystkie te czynniki mogą mieć wpływ na rozwój i aktywność biochemiczną bakterii oraz procesy związane z obiegiem węgla, azotu, fosforu, siarki i żelaza (Kuczyńska, Niewolak 2004a, 2004b). Kwasy humusowe są podatne na mikrobiologiczny rozkład i mogą wzmagać aktywność heterotroficzną bakterioplanktonu (Münster i in. 1992) oraz lipolityczną bakterii heterotroficznych (Donderski, Wódkowska 1995). Według tych ostatnich, w obecności substancji humusowych bakterie heterotroficzne mogą nabywać zdolności rozkładu substratów, które wcześniej były niedostępne. Występowanie bakterii heterotroficznych w wodzie jezior dystroficznych na obszarze WPN nie było dotychczas rozpoznane. W tym opracowaniu przedstawiono wyniki badań liczebności i składu jakościowego tych drobnoustrojów w wodzie pięciu jezior polihumusowych oraz wyniki badań eksperymentalnych dotyczących wpływu pH i rodzaju wody w pożywce na ilościowe ich oznaczenie w wodzie jeziornej.
Obiekt badań
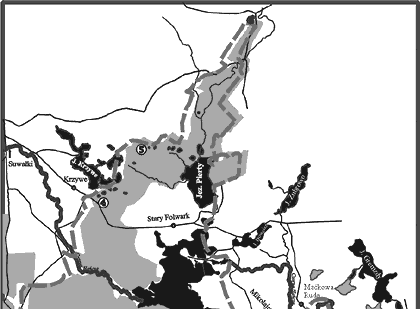
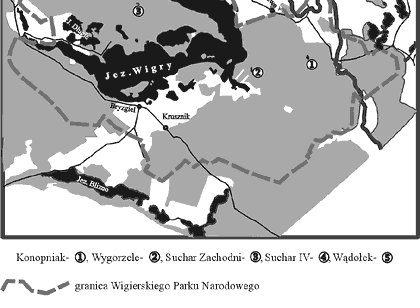
Badaniami objęto pięć jezior dystroficznych (typu „suchar”): Konopniak, Wygorzele (Ślepiec), Suchar Zachodni, Suchar IV i Wądołek. Są one rozmieszczone w północno-zachodniej (Suchar IV i Wądołek), środkowej (Suchar Zachodni) i południowo-wschodniej (Konopniak, Wygorzele) części Wigierskiego Parku Narodowego (rys. 1). Wybrane własności badanych jezior i cechy fizycznogeograficzne ich zlewni podano w tabeli 1.
Materiały i metody
Pobór prób
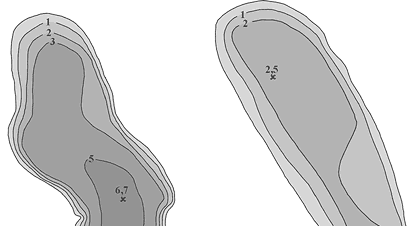
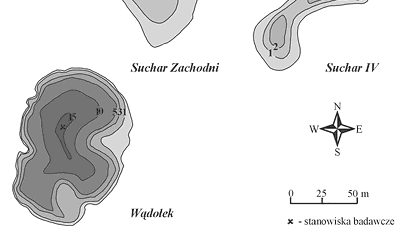
Próby wody do badań mikrobiologicznych pobierano od kwietnia 1998 roku do listopada 2000 w odstępach jednomiesięcznych, z wyjątkiem okresu zlodzenia. We wszystkich jeziorach stanowiska poboru prób znajdowały się w najgłębszym miejscu (rys. 2). Próby wody do badań pobierano na jeziorach: Konopniak, z głębokości 0,3 m, 3,4 m i 6,7 m; Wygorzele (Ślepiec), z głębokości 0,3 m, 1,3 m i 2,5 m; Suchar Zachodni, z głębokości 0,3 m, 1,2 m i 2,3 m; Suchar IV, z głębokości 0,3 m, 4,0 m i 8,0 m; Wądołek, z głębokości 0,3 m, 7,5 m i 15,0 m.
Rys. 1. Lokalizacja badanych jezior dystroficznych na obszarze Wigierskiego Parku Narodowego Powrót do poprzedniego artykułu
Próby wody powierzchniowej (z głębokości 0,3 m) pobierano bezpośrednio do jałowych naczyń szklanych z doszlifowanym korkiem, o pojemności 250 cm3; próby wody z głębszych warstw i wody przydennej (z odległości 0,3–0,5 m od dna) pobierano przyrządem Ruttnera o pojemności 2 dm3. Bezpośrednio po pobraniu wszystkie próby wody wstawiano do pojemnika (termotorby) z czynnikiem chłodzącym o temperaturze 4–6°C i przewożono do laboratorium, gdzie poddawano je badaniom mikrobiologicznym. Czas od chwili pobrania prób wody na jeziorach do wykonania analiz nie przekraczał 12 godzin.
Rys. 2. Mapy batymetryczne badanych
jezior dystroficznych Wigierskiego Parku Narodowego
Badania mikrobiologiczne
Badania mikrobiologiczne w zakresie niniejszej pracy obejmowały oznaczenia: liczebności bakterii heterotroficznych w trzech kolejnych sezonach wegetacyjnych (lata 1998–2000) w wodzie jezior: Konopniak, Wygorzele, Suchar Zachodni, Suchar IV i Wądołek; składu jakościowego (rodzajowego) tych drobnoustrojów w wodzie badanych zbiorników w 2000 roku – na podłożu z tryptonem, glukozą i ekstraktem drożdżowym (TGY 1:8) o pH 7,2, sporządzonym na wodzie destylowanej i rozcieńczonym w stosunku 1:8 (Seppänen 1971) po 7 dniach inkubacji w temperaturze 20°C; liczebności i składu jakościowego bakterii heterotroficznych w wodzie jeziora Wygorzele na podłożu TGY (1:8) sporządzonym na wodzie destylowanej i pH = 7,2 (wariant I) lub pH = 5,2 (wariant II), na wodzie jeziornej i pH = 7,2 (wariant III) lub pH = 5,2 (wariant IV), wszystkie po siedmiu dniach inkubacji w temperaturze 20°C. Identyfikację wyrosłych na tych podłożach kolonii bakterii przeprowadzano w oparciu o dane zawarte w podręcznikach Bergeya (1994), Skermana (1967) i schematów identyfikacyjnych podanych przez Hendrie’go i in. (1964). Szczepy bakteryjne do identyfikacji pobierano z kolonii różniących się morfologicznie na podłożu w płytkach Petriego. Jednocześnie zliczano oddzielnie kolonie danego typu. Na podstawie danych uzyskanych z identyfikacji każdego typu kolonii wyliczano procentowy udział poszczególnych rodzajów bakterii. Oznaczenia te wykonano w 2000 roku, w odstępach jednomiesięcznych.
Wyniki
Liczba bakterii heterotroficznych w wodzie badanych jezior, oznaczana na podłożu z tryptonem glukozą i ekstraktem drożdżowym w rozcieńczeniu 1:8 (TGY 1:8), wahała się dość znacznie (tab. 2) w zależności od zbiornika, jego głębokości i pory roku. Najmniejsze zmiany liczebności bakterii stwierdzano w wodzie powierzchniowej jeziora Konopniak (190–2580 jtk/cm3), największe w wodzie powierzchniowej jeziora Wądołek (160–28520 jtk/cm3) i jeziora Suchar IV (80–12520 jtk/1cm3). Średnio, w latach1998–2000, stwierdzano ich najmniej w wodzie jeziora Konopniak (1075–3140 jtk/cm3), najwięcej w wodzie jeziora Wądołek (2330–2980 jtk/cm3). Sezonowe zmiany liczebności bakterii heterotroficznych w poszczególnych latach charakteryzowały się maksymalnymi wartościami w różnych miesiącach, najczęściej jednak w listopadzie i sierpniu (jeziora Konopniak, Wygorzele i Wądołek), rzadziej w innych miesiącach (jeziora Suchar Zachodni i Suchar IV). Barwne szczepy bakterii heterotroficznych nie przekraczały 80 proc. ogólnej liczby tych drobnoustrojów (tab. 3). Z reguły większy procent takich szczepów stwierdzano w miesiącach wiosenno-letnich, w wodzie niektórych jezior również w październiku i listopadzie. Skład jakościowy (rodzajowy) bakterii heterotroficznych Wśród zidentyfikowanych bakterii heterotroficznych w wodzie badanych jezior dominowały rodzaje Alcaligenes (16,5–24,8 proc.), Bacillus (15,4–47,2 proc.), Flavobacterium (8–19,7 proc.) i Vibrio (8,8–25,8 proc.). Do liczniej reprezentowanych należały również bakterie z rodziny Enterobacteriaceae (2,4–13,4 proc.). Typowo wodne szczepy bakterii z rodzaju Pseudomonas stanowiły 0,5–7,2 proc. W wodzie jeziora Suchar IV liczniej reprezentowane były jeszcze bakterie z rodzaju Staphylococcus (14 proc.). Inne rodzaje bakterii reprezentowane były najczęściej w ułamkach procent (tab. 4). Wpływ rodzaju wody użytej do
sporządzania podłoża TGY (1:8) i jego pH Najmniejsze liczebności bakterii heterotroficznych w wodzie tego zbiornika oznaczano na podłożu TGY (1:8) o pH = 5,2 sporządzonym na wodzie jeziornej (wariant IV), największe na podłożu TGY (1:8) o pH = 5,2 i 7,2 sporządzonym na wodzie destylowanej (tab. 5). Wpływ rodzaju wody (destylowana, jeziorna) użytej do sporządzenia podłoża i jego pH na procent dominujących rodzajów bakterii heterotroficznych oznaczanych w wodzie z jeziora Wygorzele był niejednoznaczny (tab. 6). Na podłożu TGY (1:8) o pH = 7,2 sporządzonym na wodzie destylowanej (wariant I) uzyskiwano wyższy procent bakterii z rodzaju Flavobacterium, Micrococcus, Pseudomonas i Vibrio. Na takim samym podłożu, ale o pH 5,2 (wariant II) – więcej bakterii z rodzaju Alcaligenes, Bacillus, Micrococcus i z rodziny Enterobacteriaceae. Na podłożu TGY (1:8) o pH 7,2 sporządzonym na wodzie jeziornej (wariant III) uzyskiwano większy procent bakterii z rodzaju Alcaligenes, Bacillus i z rodziny Enterobacteriaceae. Na takim samym podłożu, ale o pH 5,2 (wariant IV) – większy procent bakterii z rodzaju Pseudomonas i niewiele więcej z rodziny Enterobacteriaceae.
Dyskusja
Liczby bakterii heterotroficznych w wodzie badanych jezior dystroficznych były na ogół dość wysokie w porównaniu z innymi jeziorami tego typu w Finlandii. Według Seppänena (1971) w wodzie jeziora humusowego Hakojärvi liczba bakterii heterotroficznych oznaczana na takim samym podłożu TGY (1:8) wahała się od kilku jkt/cm3 na przełomie marca i kwietnia (pod koniec zlodzenia) do ponad 4000 jkt/cm3 latem (w sierpniu) i wynosiła średnio od 39 do 2000 jkt/cm3. W wodzie jeziora Konopniak liczba tych bakterii była zbliżona do stwierdzonych w wodzie dystroficznego jeziora Frejen w południowej Szwecji (Schütt 1988a). Autor ten oznaczał liczbę bakterii heterotroficznych na podłożu peptonowo-kazeinowym. Donderski (1983) stwierdzał w wodzie mezotroficzno-dystroficznego Jeziora Jasne na Pojezierzu Iławskim średnio 1800 jkt/cm3 tych bakterii. Letnie maksima liczebności bakterii heterotroficznych w wodzie badanych jezior dystroficznych na obszarze WPN mogły być związane z większą dostępnością substancji organicznej. Według Lindella i in. (1995), w miesiącach letnich nie tylko temperatura wody, ale i promieniowanie słoneczne o długości fal wyższych niż UV–B wpływają na zwiększanie dostępności rozpuszczonej substancji humusowej. Nawet w naszej strefie klimatycznej do wód powierzchniowych dociera wystarczająca do fotolizy kompleksów mineralno-humusowych ilość promieni słonecznych (Wojciechowski 1999). V-Balogh i Vörös (1997) również stwierdzali większą produkcję bakterioplanktonu latem w zbiornikach bogatych w substancje humusowe. Większe liczebności bakterii heterotroficznych w wodzie powierzchniowej jeziora Wądołek mogły wynikać z większej dostępności substancji pokarmowych, powstałych na skutek działania promieni słonecznych przy wyższej temperaturze latem 1998 i 1999 roku. Natomiast większe ich liczebności stwierdzane w niektórych miesiącach wiosną, latem i jesienią 2000 roku w wodzie przydennej jeziora Wądołek mogły mieć związek z kumulacją soli mineralnych w warstwie monimolimnionu (Kuczyńska 2002). Zdanowski i in. (1992) z dopływem wód podziemnych i typem miktycznym jeziora Wądołek wiążą znaczne koncentracje żelaza w warstwie przydennej wody tego zbiornika. Najczęściej niewielki procent barwnych szczepów bakterii heterotroficznych w ogólnej liczebności tych drobnoustrojów można tłumaczyć brakiem potrzeby tworzenia barwników karotenoidowych przez te drobnoustroje w wodzie o wysokiej zawartości substancji humusowych, które ograniczają penetrację światła – widzialność krążka Secchiego w wodzie badanych jezior wynosiła 1,5–3,0 m w jeziorach Konopniak i Wądołek, 0,9–1,9 m w jeziorze Wygorzele i 1,1/1,3–2,0/2,1 m w jeziorach Suchar Zachodni i Suchar IV. W składzie jakościowym bakterii heterotroficznych w wodzie badanych jezior dystroficznych dominowały rodzaje Alcaligenes, Vibrio, Flavobacterium i Bacillus. Niektóre z nich izolował Schütt (1988a, 1988b) z wody jezior dystroficznych południowej Szwecji. Według tego autora niektóre z nich mają zdolność rozkładu monomerycznych fenoli, będących składnikiem substancji humusowych. Według Donderskiego i Burkowskiej (2000) większość bakterii heterotroficznych bytujących w wodzie jezior, ma zdolność rozkładu substancji humusowych. Ciekawym zjawiskiem jest stosunkowo niewielki udział bakterii z rodzaju Pseudomonas (0,3–7,2 proc.) i Aeromonas (0,2–1,4 proc.). Dla porównania, w wodzie oligomezotroficznego jeziora Hańcza i eutroficznego jeziora Wigry oba rodzaje bakterii należały do grupy najczęściej izolowanych (Gotkowska-Płachta 2002; Górniak 2000). Duży procent bakterii z rodzaju Bacillus (od kilkunastu procent w wodzie jeziora Konopniak i Wygorzele do około 50 proc. w wodzie jeziora Wądołek) jest cechą charakterystyczną dla jezior dystroficznych (Kuzniecow 1952, 1970). Na uwagę zasługuje również obecność w badanych próbach wody bakterii z rodzaju Enteriobacteriaceae. Ich źródłem może być zwierzyna leśna (np. bobry) i dzikie ptactwo (Kuczyńska, Niewolak 2004). W pracy tej nie stwierdzono statystycznie istotnej zależności pomiędzy liczbą bakterii heterotroficznych w wodzie badanych jezior dystroficznych a pH (3,9–6,8). Również analiza składu jakościowego bakterii heterotroficznych nie ujawniła występowania jakichś specyficznych rodzajów tych drobnoustrojów przystosowanych do życia przy niskich wartościach pH. Przeciwnie, stwierdzane rodzaje bakterii heterotroficznych są znajdowane w wielu akwenach o pH neutralnym lub alkalicznym (Donderski 1983; Nair i in. 1994). Wyniki niniejszych badań korespondują z danymi Boylena i in. (1983). Autorzy ci, badając skład jakościowy bakterii heterotroficznych w wodach powierzchniowych różniących się pH (od 4,3 do 7,0), nie stwierdzili specyficznych gatunków dominujących w wodach zakwaszonych. Według Bell i Tranvik (1993) zakwaszenie nie jest bezpośrednim czynnikiem stresującym dla bakterii, ale wywiera pośredni wpływ na procesy mikrobiologiczne w jeziorach. W badaniach eksperymentalnych przeprowadzonych w tej pracy stwierdzono, że rodzaj wody (destylowana lub jeziorna) użytej do sporządzania podłoża TGY (1:8) oraz odczyn pH (5,2 i 7,2) nie miały większego wpływu na liczebność bakterii heterotroficznych oznaczanych w wodzie jeziora Wygorzele; ilość tych bakterii w każdym wariancie doświadczenia była zbliżona. Rao i Dutka (1983) oraz Scheider i Dillon (1976) stwierdzali mniejsze liczebności bakterii heterotroficznych w wodzie jezior zakwaszonych. Dotyczyło to jednak jezior zakwaszonych eksperymentalnie bądź przez kwaśny opad, gdzie zmiany pH wody były dość gwałtowne. Historia jezior dystroficznych mierzona jest wiekiem geologicznym, a drobnoustroje występujące w wodzie takich zbiorników prawdopodobnie zaadaptowały się do niskiego pH. Na liczebność bakterii heterotroficznych w wodzie badanych jezior dystroficznych większy wpływ ma zapewne zawartość substancji odżywczych i soli mineralnych (zawartość azotu ogólnego wynosiła 0,71–2,91 mg/dm3, fosforu ogólnego 0,063–0,341 mg/dm3) i prawdopodobnie związków humusowych (niebadanych w tej pracy). Z literatury wiadomo (Donderski, Wódkowska 1995; Banz i in. 1998; Coates i in. 2000; Donderski, Burkowska 2000; Elo i in. 2000; Esham i in. 2000), że wiele bakterii heterotroficznych występujących w wodach powierzchniowych, posiada zdolność degradacji takich związków. Zwraca uwagę dość znaczny procent bakterii z rodzaju Bacillus oznaczanych na podłożu TGY (1:8) o pH = 5,2 sporządzonym na wodzie destylowanej i jeziornej (wariant II i IV, odpowiednio 37,6 i 25,7 proc.) oraz na takim samym podłożu o pH = 7,2 sporządzonym na wodzie jeziornej (wariant III, 30,0 proc.) przy względnie niższym udziale tych bakterii na podłożu z wodą destylowaną (wariant I – 13,4 proc.). Skądinąd większy procent bakterii z rodzaju Pseudomonas i Arthrobacter, oznaczanych na podłożu TGY (1:8) o pH 5,2 sporządzonym na wodzie jeziornej (wariant IV), nie daje jednoznacznej odpowiedzi dotyczącej wpływu pH na rozwój badanych bakterii. Z literatury wiadomo, że bakterie z rodzaju Arthrobacter izolowane z wody jezior humusowych (Tranvik 1992) posiadały zdolność do wzrostu i wykorzystywania kwasów fulwowych, co wydawałoby się potwierdzać największy udział tego rodzaju bakterii na podłożu TGY (1:8) o pH = 5,2 sporządzonym na wodzie jeziornej (wariant IV).
Wnioski
1. Liczba bakterii heterotroficznych oznaczanych na podłożu z tryptonem, glukozą i ekstraktem drożdżowym w rozcieńczeniu 1:8 (TGY 1:8) w wodzie badanych jezior dystroficznych zmieniała się w szerokich granicach, zależnie od głębokości i pory roku; najmniejsze zmiany ich liczebności występowały w wodzie jeziora Konopniak (190–2580 jkt/cm3), największe w wodzie jeziora Wądołek (160–28520 jkt/cm3) i w wodzie przydennej jeziora Suchar IV (50–12520 jkt/cm3). 2. Sezonowe zmiany liczebności bakterii heterotroficznych charakteryzowały się maksymalnymi wartościami w lipcu i/lub w sierpniu (jeziora Konopniak, Wygorzele i Wądołek), okresowo również wiosną (jezioro Suchar IV) lub jesienią (jezioro Suchar Zachodni). 3. Letnie maksima liczebności bakterii heterotroficznych w wodzie badanych jezior dystroficznych mogą mieć związek ze zwiększeniem dostępności substancji organicznej oraz wzrostem aktywności metabolicznej drobnoustrojów przy wyższej temperaturze wody. Liczniejsze zaś występowanie tych bakterii w wodzie w powierzchniowej mogło być związane z większym nagromadzeniem substancji organicznej w tej warstwie wody. W jeziorach dystroficznych mutagenny wpływ światła słonecznego może być niwelowany dzięki wysokiej zawartości substancji humusowych w wodzie, ograniczającej penetrację promieni świetlnych w zakresie UV. Świadczy o tym niewielki procent bakterii wytwarzających barwniki karotenoidowe w ogólnej liczbie bakterii heterotroficznych. 4. W składzie jakościowym bakterii heterotroficznych w wodzie badanych jezior dystroficznych dominującymi były bakterie z rodzaju Alcaligenes (16,5–24,8 proc.), Bacillus (13,4–47,2 proc.), Flavobacterium (8,0–19,7 proc.) i Vibrio (8,8–25,8 proc.), znane ze zdolności do degradacji monomerycznych fenoli będących składnikami substancji humusowych. 5. Wyizolowane bakterie heterotroficzne występują powszechnie w większości akwenów słodkowodnych. Wysoki udział bakterii z rodzaju Bacillus potwierdza typowe dla bakterii przetrwalnikujących uwarunkowania do życia w wodach ubogich w łatwo rozkładalną substancję organiczną. 6. W badaniach eksperymentalnych nie stwierdzono istotnego związku między pH (5,2 i 7,2) i rodzajem wody (destylowana, jeziorna) użytej do sporządzania podłoża TGY (1:8) a liczbą bakterii heterotroficznych oznaczanych w wodzie z jeziora Wygorzele. Liczba tych drobnoustrojów w każdym wariancie doświadczenia była zbliżona. Może to sugerować, iż głównym czynnikiem determinującym liczebność bakterii heterotroficznych w badanych jeziorach jest dostępność substancji organicznej.
Podsumowanie
Badano liczebność i skład jakościowy bakterii heterotroficznych w wodzie jezior dystroficznych: Konopniak, Wygorzele, Suchar Zachodni, Suchar IV i Wądołek na terenie Wigierskiego Parku Narodowego. Badania prowadzono w trzech kolejnych sezonach wegetacyjnych, w latach 1998–2000. Dodatkowo w 2000 roku sprawdzano eksperymentalnie wpływ pH (5,2 i 7,2) i rodzaju wody (destylowana, jeziorna) użytej do sporządzania podłoża TGY (1:8) na liczbę i skład jakościowy bakterii oznaczanych w wodzie z jeziora Wygorzele, o widzialności krążka Secchiego 0,9–1,9 m, temperaturze mniej więcej jednakowej od powierzchni do dna i pH około 5,0. Liczba bakterii heterotroficznych w wodzie badanych jezior dystroficznych w okresie badawczym wahała się w zakresie 2–3 rzędów wielkości, zależnie od głębokości, pory roku i okresu badań. Częstokroć więcej ich stwierdzano w powierzchniowej warstwie wody (z głębokości 0,3 m) i latem, rzadziej wiosną i/lub jesienią. Wśród bakterii heterotroficznych zidentyfikowanych w 2000 roku dominowały rodzaje: Alcaligenes, Bacillus, Flavobacterium i Vibrio. W badaniach eksperymentalnych nie stwierdzono wyraźnego związku pomiędzy pH czy rodzajem wody użytej do sporządzania podłoża (TGY 1:8) a liczbą i składem jakościowym bakterii heterotroficznych oznaczanych w wodzie z jeziora Wygorzele.
Abstract
The occurrence of heterotrophic bacteria in dystrophic lakes’ water on the area of Wigry National Park Studies on the number and composition of heterotrophic bacteria in waters of 5 dystrophic lakes: Konopniak, Wygorzele Wygorzele (Ślepiec), Suchar Zachodni, Suchar IV, Wądołek located on the area of Wigry National Park were carried out in three annual cycles, in 1998-2000. Additionally, in 2000, effect of pH (5,2 and 7,2) and kind of water (dystrophic lakes’ water and distilled water) used to preparation of TGY (1:8) medium on the number and qualitative composition of heterotrophic bacteria from Wygorzele lake (visibility of Secchi disk 0,9–1,9 m, almost the same water temperature form surface to the bottom, and pH about 5,0) was tested. The number of heterotrophic bacteria ranged within 2-3 orders of magnitude, depending on depth of the lake, season and the investigation period. The most numerous heterotrophic bacteria were detected in surface water (0,3 m) and more often in summer than in autumn or spring. Among heterotrophic bacteria identified in 2000, Alcaligenes, Bacillus, Flavobacterium and Vibrio genera predominated. pH and kind of water used to preparation of TGY (1:8) medium did not considerably effect quantity or qualitative composition of heterotrophic bacteria from Wygorzele lake.
Tabela 1. Wybrane właściwości morfometryczne i hydrologiczne badanych jezior dystroficznych na obszarze Wigierskiego Parku Narodowego (według Bajkiewicz-Grabowskiej i in. 1992)
Powrót do poprzedniego artykułu
Tabela 2. Ogólna liczba bakterii heterotroficznych oznaczana na podłożu TGY (1:8) w wodzie jezior dystroficznych na obszarze Wigierskiego Parku Narodowego w latach 1998–2000 (wartości średnie z 3 równoległych oznaczeń w 1 cm3 wody)
* – głębokość poboru prób wody
Tabela 3. Procentowy udział barwnych bakterii heterotroficznych w ogólnej liczbie tych drobnoustrojów oznaczanych na podłożu TGY (1:8) w jeziorach dystroficznych na obszarze Wigierskiego Parku Narodowego w latach 1998–2000 (wartości średnie z 3 równoległych oznaczeń w 1 cm3 wody)
Tabela 4. Skład jakościowy i procentowy udział różnych rodzajów bakterii heterotroficznych w jeziorach dystroficznych na podłożu TGY (1:8) sporządzonym na wodzie destylowanej pH 7,2 w 2000 roku (średnie wartości z poszczególnych miesięcy w całym roku)
Tabela 5. Liczba bakterii heterotroficznych w wodzie jeziora Wygorzele (Ślepiec) w roku 2000 oznaczanych na podłożu TGY (1:8), sporządzonym na wodzie jeziornej i destylowanej (pH 5,2 i 7,2)
Tabela 6. Skład jakościowy i procentowy udział różnych rodzajów bakterii heterotroficznych oznaczanych na podłożu TGY (1:8), sporządzonym na wodzie jeziornej i destylowanej (pH 5,2 i 7,2) w jeziorze Wygorzele (Ślepiec) w roku 2000 (średnie wartości z poszczególnych miesięcy w całym roku)
Literatura
Bajkiewicz-Grabowska E., 1992: Charakterystyka fizycznogeograficzna i klimatyczna parku. W: Jeziora Wigierskiego Parku Narodowego. Stan eutrofizacji i kierunki ochrony. Wrocław, Zeszyty Naukowe PAN „Człowiek i Środowisko”, s. 11–17. Bell R. T., Tranvik L., 1993: Imapct of Acidification and Liming on the Microbial Ecology of Lakes. Ambio, 22 (5):325–330. Benz M., Schink B., Brune A., 1998: Humic Acid Reduction by Propionibacterium freudenreichii and Other Fermenting Bacteria. Appl. Environ. Microbiol., 64 (11):4507–4512. Bergey’s Manual of Determinative Bacteriology. 1994. Red. Holt J. G., Krieg N. R., Sneath P. H. A., Staley J. T., Williams S. T., The Williams and Wilkins Company, USA. Boylen C. W., Shick M. O., Roberts D. A., Singer R., 1983: Microbiological Survey of Adirondack Lakes with Various pH Values. Appl. Environ. Microbiol., 45:1538–1544. Coates J. D., Ellis D. J., Blunt-Harris E. L., Gaw C. V., Roden E. E., Lovley D .R., 1998: Recovery of Humic-Reducing Bacteria from a Diversity of Environments. Appl. Environ. Microbiol., 64 (4):1504–1509. Donderski W., 1983: Tlenowe bakterie heterotroficzne jezior o różnej trofii. Rozpr. UMK, Toruń. Donderski W., Burkowska A., 2000: Metabolic Activity of Heterotrophic Bacteria in the Presence of Humic Substances and Their Fractions. Pol. J. Environ. Stud., 9 (4):267–271. Donderski W., Wódkowska A., 1997: Humic Substances as Source of Carbon and Nitrogen for Heterotrophic Bacteria Isolated from Lakes of Different Trophy. Pol. J. Environ. Stud., 6 (4):9–15. Elo S., Maunuksela L., Salkinoja-Salonen M., Smolander A., Haahtela K., 2000: Humus bacteria of Norway spruce stands: plant growth promoting properties and birch, red fescue and alder colonizing capacity. FEMS Microbiol. Ecol., 31 (2):143–152. Gotkowska-Płachta A., 2002: Studium mikrobiologiczne wód jeziora Hańcza. Olsztyn, Uniw. Warm.-Maz., rozprawa doktorska. Górniak D., 2000: Struktura i funkcja bakterioplanktonu systemu rzeczno-jeziornego rzeki Czarnej Hańczy. Olsztyn, Uniw. Warm.-Maz., rozprawa doktorska. Hendrie M. S., Hodgkiss W., Shewan J. M., 1964: Considerations on organisms of the Achromobacter-Alcaligenes Group. Annales de L’Institute Pasteur de Lille, 15:43–59. Kuczyńska A., 2002: Mikrobiologiczna charakterystyka wybranych jezior dystroficznych na terenie Wigierskiego Parku Narodowego. Olsztyn, Uniw. Warm.-Maz., rozprawa doktorska. Kuczyńska A., Niewolak S., 2004a: Ocena stopnia zanieczyszczenia i stanu sanitarno-bakteriologicznego wody jezior dystroficznych na obszarze Wigierskiego Parku Narodowego, maszynopis. Kuczyńska A., Niewolak S., 2004b: Sezonowe zmiany liczebności bakterii czynnych w obiegu azotu w wodzie jezior dystroficznych na obszarze Wigierskiego Parku Narodowego, maszynopis. Kuzniecow S. I., 1952: Rol mikroorganizmow w krugoworotie wieszcziestwa w ozierach. Moskwa–Leningrad, AN SSSR. Kuzniecow S. I., 1970: Mikroflora ozier i jejo geochimiczieskaja diejatielnost’. Leningrad. Lindell M. J., Granéli W. H., Tranvik L. J., 1995: Enhanced bacterial growth in response to photochemical transformation of dissolved organic matter. Limnol. Oceanogr., 40:195–199. Münster U., Numinen J., Einio P., Overbeck J., 1992: Extracellular enzymes in a small polyhumic lake: origin, distribution and activities. Hydrobiol., 243/244:47–59. Nair S., Loka Bharathi P. A., Chandramohan D., 1994: Culturable heterotrophic bacteria from the euphotic zone of the Indian Ocean during the summer monsoon. Ocean. Act., 17 (1):63–68. Rao S. S., Paolini D., Leppard C. G., 1984: Effects of low-pH stress on the morphology and activity of bacteria from lakes receiving acid precipitation. Hydrobiol., 114:115–121. Scheider W., Dillon P. J., 1976: Neutralization and fertilization of acidified lakes near Sudbury, Ontario. Wat. Poll. Res. Can., 11:93–100. Schütt Ch., 1988a: XII. Ecology of Aquatic Organisms. 1. Microbes (continued). Habitat-specific plasmid patterns in natural bacterial populations of fresh-water lakes. Verh. Internat. Verein. Limnol., 23:1821– 1824. Schütt Ch., 1988b: Plasmid-DNA in natural bacterial populations of four brownwater lakes (South Sweden). Arch. Hydrobiol. Beih., 31:133–139. Seppänen H., 1971: Bacteriological studies on waters with low electrolyte concentration. Otaniemi, Helsinki University of Technology Research Papers, 34:1–87. Skerman V. B. D., 1967: A guide to the identification of the genera of bacteria with methods and digest of generic characteristics. Baltimore, Williams and Wilkins Comp. Stangenberg M., 1936: Szkic limnologiczny na tle stosunków hydrochemicznych Pojezierza Suwalskiego. Warszawa, Inst. Bad. Lasów Państwowych, Seria A, Rozpr. 1, spraw. nr 19, s. 85, il., tab., bibliogr. 7–54. Tranvik L. J., 1992: Allochthonous dissolved organic matter as an energy source for pelagic bacteria and the concept of the microbial loop. Hydrobiol., 229:107–114. V.-Balogh K., Vörös L., 1997: High bacterial production in hypertrophic shallow reservoirs rich in humic substances. Hydrobiol., 342/343:63–70. Wojciechowski I., 1999: Warunki funkcjonowania ekosystemów torfowiskowych i wodno-torfowiskowych w Polsce. Problemy aktywnej ochrony ekosystemów wodnych i torfowiskowych w polskich parkach narodowych. Red. Radwan S., Kornijow R. Lublin, Wyd. Uniwersyt. M. Curie-Składowskiej, s. 57–65. Zdanowski B., 1999: Eutrofizacja jezior Wigierskiego Parku Narodowego: zagrożenia i ocena. W: Zdanowski B., Kamiński M., Martyniak A. (red.): Funkcjonowanie i ochrona ekosystemów wodnych na obszarach chronionych. Olsztyn, IRS. Zdanowski B., Karpiński A., Prusik S., 1992: Warunki środowiskowe wód jezior Wigierskiego Parku Narodowego. W: Jeziora Wigierskiego Parku Narodowego. Stan eutrofizacji i kierunki ochrony. Wrocław, Zeszyty Naukowe PAN „Człowiek i Środowisko”, s. 35–62.
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||