|
AUGUSTOWSKO-SUWALSKIE TOWARZYSTWO NAUKOWE
Proszę chwilę zaczekać, ładuję stronę ... |
|
AUGUSTOWSKO-SUWALSKIE TOWARZYSTWO NAUKOWE
Proszę chwilę zaczekać, ładuję stronę ... |
|
Maciej Karpowicz, Andrzej Górniak, Adam Cudowski
Struktura zespołów zooplanktonu
skorupiakowego
Wstęp Jezioro Wigry ma ponad 100-letnią historię badań zooplanktonu, zapoczątkowanych przez B. Heynemana w 1902 roku i kontynuowanych przez pracowników wigierskiej Stacji Hydrobiologicznej, pod kierunkiem Alfreda Lityńskiego w latach 1920–1939. Późniejsze badania były prowadzone w latach 1963–1967, w ramach sporządzania operatu rybackiego zagospodarowania jezior (Leszczyńska-Kowalska 1992), następnie w latach 1981–1995 przez Zakład Hydrobiologii Instytutu Ekologii Polskiej Akademii Nauk i Instytutu Rybactwa Śródlądowego (Karabin, Ejsmont-Karabin 1999) oraz na początku XXI wieku przez Zakład Hydrobiologii Uniwersytetu w Białymstoku (Górniak i in. 2002). Niniejsze opracowanie prezentuje aktualną strukturę zespołów zooplanktonu skorupiakowego jeziora Wigry na tle ponad 100 lat obserwacji oraz ich przestrzenne zróżnicowanie na tle trofii wód.
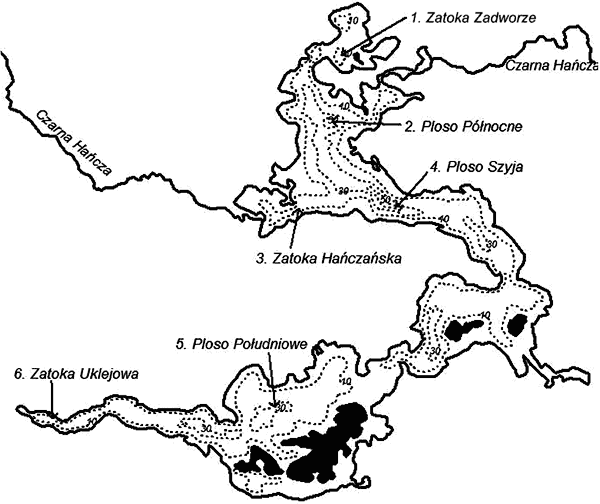
Teren i metody Wigry to jezioro rynnowo-wytopiskowe, otoczone od północy, zachodu i wschodu osadami morenowymi, a od południa równiną sandrową (Ber 1981). Rozległa powierzchnia 2170 ha i głębokość maksymalna do 73 metrów sprawiają, że jest to jeden z największych rezerwuarów wody północno-wschodniej Polski (Górniak red. 2006). Posiada rozwiniętą linię brzegową z licznymi półwyspami, zatokami i wyspami, a część tych ostatnich powstaje na skutek depozycji wytrącającego się kalcytu w trakcie biogenicznej demineralizacji wód. Dzięki temu Wigry nie posiadają jednolitego charakteru biologicznego, co wpływa na zadziwiającą różnorodność przyrody oraz bogactwo gatunkowe fauny i flory. Badania terenowe prowadzono w pierwszej dekadzie sierpnia 2007 roku, w warunkach suszy hydrologicznej, na sześciu stanowiskach (rys. 1). Próby zooplanktonu pobierano 5-litrowym czerpaczem Bernatowicza z sześciu stanowisk, reprezentujących odmienne części jeziora (rys. 1), oddzielnie dla każdej warstwy wody: epilimnionu, metalimnionu i hypolimnionu. Próby zagęszczono filtrując je przez siatkę planktonową o średnicy oczek ok. 30 µm, a następnie utrwalono 4% formaldehydem. Po sedymentacji dokonywano identyfikacji taksonomicznej planktonu skorupiakowego, określano jego liczebność, a biomasę przedstawicieli poszczególnych gatunków określono na podstawie zależności długość/ciężar według Balushkina i Vinberga (1979). Na podstawie struktury zooplanktonu skorupiakowego wyliczono zooplanktonowe wskaźniki trofii (WST) wg Karabina i Ejsmont-Karabin (1985). Pomiary i analizy chemiczne wód, wykonane według metod i procedur opisanych przez Górniaka (red. 2006), posłużyły do wyliczenia wskaźników żyzności według Carlsona (1977) oraz Kratzer i Brezonik (1981).
Rys. 1. Mapa jeziora Wigry z rozmieszczeniem stanowisk poboru prób
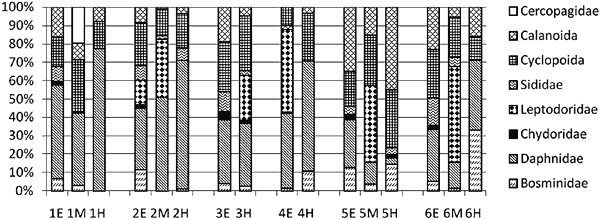
Aktualna struktura zespołów zooplanktonu skorupiakowego Pelagial jeziora Wigry aktualnie jest zasiedlony przez bogaty gatunkowo zespół zooplanktonu skorupiakowego. Stwierdzono występowanie 24 gatunków Crustacea (tab.1), w tym 11 Cladocera, 5 Calanoida i 8 Cyclopoida. Na wszystkich stanowiskach było od 14 do 17 gatunków, a różnice dotyczyły wyłącznie organizmów o najmniejszej liczebności. Widoczne były różnice obfitości i dominacji poszczególnych gatunków skorupiaków w różnych częściach jeziora oraz w słupie wody.
Tabela 1. Udział procentowy poszczególnych gatunków w biomasie zooplanktonu skorupiakowego na badanych stanowiskach w jeziorze Wigry latem 2007 roku
(+ oznacza udział poniżej
jednego procenta; E – epilimnion; M –
metalimnion;
Wśród wioślarek w liczebności i biomasie dominowała Daphnia cucullata Sars, która występowała na każdym stanowisku w całym słupie wody, a w epilimnionie stanowiła 24–50% biomasy całego zooplanktonu skorupiakowego. Wszystkie osobniki tego gatunku posiadały wydłużony hełm, wiązany z dymorfizmem sezonowym. Cechą charakterystyczną jeziora Wigry jest również współwystępowanie wielu gatunków rodzaju Daphnia. Na każdym stanowisku występowały 3–4 gatunki tego rodzaju, przy czym obecność D. hyalina i D. longispina stwierdzano jedynie w chłodnym hypolimnionie. Wartym podkreślenia jest występowanie na wszystkich stanowiskach D. cristata, która jest gatunkiem typowym dla jezior o niskiej trofii. Na każdym badanym stanowisku występował duży pelagiczny drapieżnik Leptodora kindtii. Ponadto w Zatoce Zadworze stwierdzono Bythotrephes longimanus – palearktyczny gatunek, który preferuje większe, głębokie jeziora z wychłodzonym hypolimnionem (MacIsaac i in. 2000). W Zatokach Uklejowej i Hańczańskiej występowało więcej gatunków typowych dla wysokiej trofii: Chydorus sphaericus i Diaphanosoma brachyurum. Rząd Calanoida reprezentowany był przez pięć gatunków. Wyraźnym dominantem był Eudiaptomus gracilis, który stanowił 75–80% liczebności Calanoida. Stwierdzono również występowanie dwóch rzadkich już dla Polski skorupiaków: Heterocope appendiculata i Eurytermora lacustris. Są to gatunki chłodnolubne, pochodzenia bałtoskandynawskiego, przy czym E. lacustris jest uważany za relikt morski (Brzęk 1988). W 2007 roku spotkano nowy dla fauny Wigier gatunek – Acantodiaptomus denticornis Wierzejski, który jest gatunkiem eurytermicznym występującym w dużych jeziorach, szczególnie często w górskich, lecz bywa spotykany również w stawach (Rybak 1994). Oczliki (Cyclopoida) stanowiły jedynie 9–27% biomasy całego zooplanktonu skorupiakowego. Przy stosunkowo niewielkiej liczebności, stwierdzono duże zróżnicowanie gatunkowe (osiem gatunków). O biomasie zespołu decydował Mesocyclops leuckarti, który stanowił 80% liczebności wszystkich oczlików. W 2007 roku zaobserwowano również wystąpienie nowego dla Wigier gatunku Metacyclops gracilis, którego obecność stwierdzono na wszystkich stanowiskach. Liczebność dorosłych skorupiaków w epilimnionie jeziora Wigry wahała się w przedziale 70–234 os./dm3. Najmniejsze liczebności obserwowano w Plosie Szyja i w Plosie Południowym. Większa liczebność zooplanktonu skorupiakowego wystąpiła w Plosie Północnym oraz w badanych zatokach – ponad 200 os./dm3 (rys. 2). Na uwagę zasługuje również duża liczebność form larwalnych (nauplii) Cyclopoida – ponad 190 os./dm3 w Zatoce Uklejowej.
Rys. 2. Liczebność (os./dm3) oraz struktura
Crustacea jeziora Wigry latem 2007 roku;
Biomasa zooplanktonu w epilimnionie wahała się od 3,8 mg św.m./dm3 w Plosie Południowym do 6,6 mg św.m./dm3 w Zatoce Zadworze. Niską wartością odznaczało się Ploso Szyja, a wyższe wartości występowały w zatokach. Wysoka biomasa zooplanktonu skorupiakowego spowodowana jest dominacją dużych gatunków Crustacea. Wpływ na to mogły mieć prowadzone działania biomanipulacyjne, polegające na zwiększaniu liczebności ryb drapieżnych (szczupak), które zmniejszają liczebność ryb karpiowatych. Skutkiem tego wystąpiło zmniejszanie presji na duży zooplankton skorupiakowy, który w dalszej kolejności ogranicza również zakwity glonów planktonowych przyczyniające się do powstania okresowych deficytów tlenu w profundalu. Procentowy udział biomasy poszczególnych taksonów w strukturze zooplanktonu został przedstawiony na rys. 3 i posłużył do wyliczenia wskaźnika stanu trofii (WST) jeziora.
Rys. 3. Struktura biomasy letniego zooplanktonu skorupiakowego
jeziora Wigry
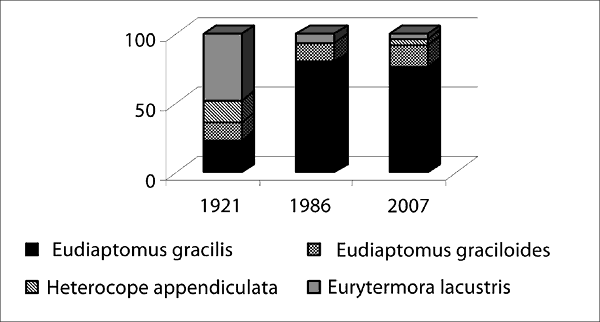
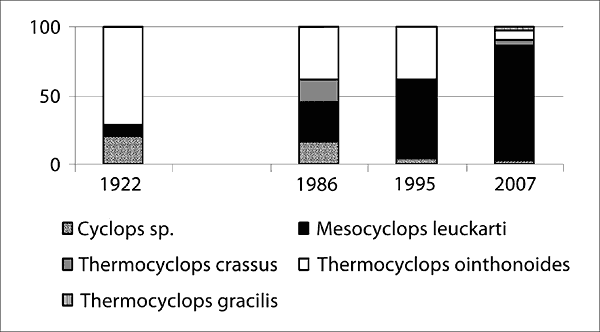
Wieloletnie zmiany struktury zooplanktonu jako wskaźnika stanu limnologicznego jeziora Skład gatunkowy zooplanktonu jeziora Wigry w pierwszej połowie XX wieku był typowy dla dużych zbiorników mezotroficznych (Karabin, Ejsmont-Karabin 1999). Analizy przeprowadzone w latach 1981–1986 wykazały jednak przekształcenia struktury zespołów zooplanktonu i świadczyły o postępującej powolnej eutrofizacji jeziora. Wyrażało się to m.in. wzrostem liczby gatunków Cyclopoida oraz zwiększającym się udziałem tej grupy w biomasie skorupiaków planktonowych, spadkiem znaczenia gatunków typowych dla jezior o niskiej trofii, przy jednoczesnym wzroście roli organizmów charakterystycznych dla eutrofii. Najbardziej radykalnej przebudowie uległ zespół Calanoida. W latach dwudziestych ponad połowę liczebności tego taksonu stanowiły gatunki typowe dla mezotrofii – Heterocope appendiculata i Eurytemora lacustris. Dzisiaj te gatunki występowały sporadycznie, a wyraźna była dominacja Eudiaptomus gracilis, który stanowił 80–90 % liczebności wszystkich Calanoida. W rzędzie Cyclopoida w latach osiemdziesiątych obserwowano wzrost liczby gatunków oraz zwiększający się udział tej grupy w biomasie skorupiaków planktonowych, co świadczyło o postępującej eutrofizacji jeziora. Notowano również istotną odmienność północnej i południowej części jeziora, gdy chodzi o obfitość i strukturę zespołu Cyclopoida. W części północnej i środkowej zespół ten charakteryzował się dość wysoką biomasą (19–79%), podczas gdy w części południowej stanowił jedynie 12–15% (Karabin, Ejsmont-Karabin 1991). Fakt ilościowego zróżnicowania zespołów skorupiaków w poszczególnych partiach jeziora zauważył już na początku wieku Lityński, który stwierdził: „Na Plosie Północnym uderza rzadsze występowanie wioślarek oraz obfitszy udział w planktonie widłonogów Cyclopidae”. Od pewnego czasu notuje się jednak zmianę tej sytuacji i już w 2002 roku Cladocera przeważały nad Cyclopoida. Obecnie oczliki stanowią 10–27% biomasy zooplanktonu skorupiakowego. Wskazuje to na poprawę warunków panujących w części północnej akwenu. W strukturze gatunkowej zespołu na przestrzeni ostatnich stu lat obserwuje się spadek znaczenia Thermocyclops oithonoides oraz Cyclops scutifer na rzecz wzrastającej dominacji Mesocyclops leuckarti. Obecnie Cyclops scutifer występował jedynie na głębszych stanowiskach. Pojawiły się natomiast nowe gatunki: Thermocyclops crassus, Metacyclops gracilis oraz przedstawiciele rodzaju Microcyclops.
Rys. 4. Wieloletnie zmiany struktury w zespołach Calanoida i Cyclopoida
W rzędzie Cladocera zaszły najmniejsze zmiany. Zwiększało się bogactwo gatunkowe, przy jednoczesnym wzroście znaczenia D. cucullata, a spadku E. crassicornis. Jedynie w Zatoce Zadworze utrzymywała się równowaga liczebności tych dwóch gatunków. Analizy zooplanktonu skorupiakowego przeprowadzone w ostatnich latach (1995–2007) skłaniają do bardziej optymistycznych wniosków i wykazują spowolnienie tempa eutrofizacji jeziora Wigry. Wyraża się to zmniejszaniem znaczenia Cyclopoida oraz występowaniem gatunków charakterystycznych dla niskiej trofii. Struktura zooplanktonu w 2007 roku wykazuje dalszą poprawę warunków w części północnej jeziora, przy wzroście żyzności w części południowej, wyrażonej zwiększaniem dominacji Cyclopoida oraz gatunków typowych dla eutrofii: Chydorus spaearicus oraz Diaphanosoma brachyurum.
Zooplanktonowe wskaźniki trofii jeziora Struktura zespołów zooplanktonu skorupiakowego zasiedlającego epilimnion w szczycie stagnacji letniej posłużyła również do wyznaczenia wskaźników trofii jeziora z wykorzystaniem wzorów regresji Karabina i Ejsmon-Karabin (1985). Wskaźniki opierające się na strukturze zooplanktonu są doskonałym uzupełnieniem dla TSI (Trophic State Index) Carlsona (1977), gdyż ukazują reakcję wyższego poziomu troficznego na zmiany zachodzące w jeziorze. Już w latach dwudziestych stwierdzono znaczne zróżnicowanie ilościowe zespołu zooplanktonu skorupiakowego pomiędzy poszczególnymi częściami zbiornika, świadczące o zróżnicowaniu warunków panujących w zbiorniku. Różnice między stanowiskami, gdy chodzi o ich żyzność, najlepiej można zilustrować używając planktonowych wskaźników trofii (Karabin, Ejsmont-Karabin, 1985). W niniejszej pracy posłużono się trzema wskaźnikami dotyczącymi Crustacea.
Tabela 2. Zooplanktonowe
wskaźniki stanu trofii w epilimnionie jeziora Wigry
Pierwsze dwa wskaźniki dotyczą udziału biomasy Cyclopoida w biomasie całkowitej zooplanktonu skorupiakowego (CCB) oraz stosunku biomasy Cyclopoida do Cladocera (BR). W 1986 roku w północnej i środkowej części jeziora Wigry Cyclopoida stanowiły 19–79% biomasy zooplanktonu skorupiakowego, gdy w części południowej wartość tego wskaźnika nie przekraczała 15%. Od roku 2002 widoczna jest przewaga Cladocera nad Cyclopoida (Rudź, Górniak 2002). Analizy przeprowadzone w 2007 roku wykazały dalsze zmniejszenie dominacji Cyclopoida w północnej części jeziora, co świadczy o zahamowaniu procesu eutrofizacji. W części południowej zaobserwowano natomiast tendencję wzrostu znaczenia Cyclopoida, szczególnie w Zatoce Uklejowej, gdzie oprócz wzrostu znaczenia taksonu, zaobserwowano prawie czterokrotnie większą liczebność form larwalnych (nauplii) nad formami dorosłymi, co może wskazywać na jeszcze większą dominację tego rzędu i świadczyć o wzroście żyzności. Następny wskaźnik informuje o udziale grupy gatunków wskaźnikowych wysokiej trofii w liczebności wszystkich gatunków wskaźnikowych (IS). W tym celu wyodrębniono dwie grupy zooplanktonu skorupiakowego. Pierwszą, charakterystyczną dla jezior o niższej trofii, tworzą: Daphnia cucullata, D. cristata oraz Heterocope appendiculata. W skład drugiej grupy ekologicznej, której optimum warunków środowiskowych występuje w zbiornikach o wyższej trofii, wchodzą: Mesocyclops leuckarti, Thermocyclops oithonoides, Diaphanosoma brachyurum i Chydorus sphaericus (Karabin, Ejsmont-Karabin 1994). Obserwowano podobne jak w poprzednich wskaźnikach zróżnicowanie, ale różnice pomiędzy poszczególnymi stanowiskami były niewielkie. Struktura zooplanktonu wskazywała na panujące w zbiorniku warunki mezoeutroficzne. Jedynie Ploso Szyja charakteryzowało się mniejszą żyznością, bliską oligotrofii. Widoczne były zmiany w strukturze i dominacji poszczególnych taksonów zooplanktonu, świadczące o poprawie warunków w północnej części jeziora Wigry, przy jednocześnie postępującej eutrofizacji części południowej.
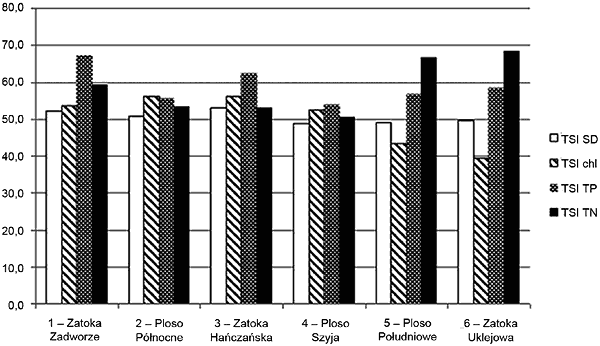
Fizykochemiczne wskaźniki stanu trofii Analizy fizykochemiczne wody posłużyły do ustalenia żyzności poszczególnych części akwenu oraz porównania z zooplanktonowymi wskaźnikami trofii. Wykorzystano powszechnie stosowany numeryczny zbiór Wskaźników Stanu Trofii (ang. TSI – Trophic State Index) opracowany przez Carlsona (1977). Wskaźniki te obliczono na podstawie: zawartości chlorofilu „a” (TSIchl), widzialności krążka Secchi’ego (TSISD), stężenia fosforu całkowitego (TSITP). Wykorzystano również TSI wyliczony na podstawie stężenia azotu całkowitego (TSITN) (Kratzer i Brezonik 1981).
Tabela 3. Współczynniki TSI na podstawie parametrów fizykochemicznych wody
Model Carlsona opiera się na założeniu, że pomiędzy letnim stężeniem fosforu całkowitego, widzialnością krążka Secchi’ego i stężeniem chlorofilu „a”, istnieje zależność pozwalająca na porównywalność obliczonych wskaźników. Kompensacja i zrównoważenie wszystkich wskaźników są charakterystyczne dla jezior o rozwoju harmonijnym i niewielkim stopniu przekształceń antropogenicznych. Jednak współcześnie w wielu jeziorach na Pojezierzu Mazurskim oraz na terenie całej Polski obserwowany jest brak kompensacji TSI, a zazwyczaj wyższe w stosunku do innych wartości TSITP traktowane są jako wyróżnik podstawowy (Hillbricht-Ilkowska i Wiśniewski 1994). Taka sytuacja zaistniała w południowej części jeziora Wigry, gdzie obserwowano wyższe wartości TSITP oraz TSITN, co wskazuje na większą podatność na eutrofizację. Równowaga i kompensacja poszczególnych wskaźników widoczne są w rynnowych częściach jeziora (Ploso Północne i Szyja) oraz w Zatoce Hańczańskiej. Widoczne jest zmniejszenie stężenia biogenów w części północnej i ich kompensacja z pozostałymi wskaźnikami, co wskazuje na stabilizowanie się warunków i zahamowanie procesu eutrofizacji tej części jeziora.
Rys. 5. Zróżnicowanie poszczególnych wskaźników TSI na badanych stanowiskach
Najbardziej istotny wpływ na obniżenie poziomu trofii Wigier miała modernizacja oczyszczalni ścieków w Suwałkach przeprowadzona w 1996 roku, która usprawniła procesy defosfatacji, nitryfikacji i denitryfikacji (Zdanowski 1999). Pomimo to, północna część jeziora nadal wykazuje nieco wyższą produktywność niż część południowa, ale ta część jeziora już w latach dwudziestych wykazywała cechy świadczące o wyższej trofii, w porównaniu do pozostałych stanowisk.
Wnioski Jezioro Wigry utrzymuje wysoką różnorodność zooplanktonu skorupiakowego zasobnego w 24 gatunki, dzięki zróżnicowaniu pionowemu środowiska, które stwarza dodatkowe nisze ekologiczne i umożliwia występowanie dużej liczby gatunków. Współwystępowanie jest szczególnie widoczne w przypadku rodzaju Daphnia, gdzie na każdym stanowisku występowały 3–4 gatunki. Do szczególnych walorów jeziora należy zaliczyć obecność rzadkich już dla wód Polski gatunków: Eurytermora lacustris, Heterocope appendiculata, Bythotrephes longimanus. Struktura zooplanktonu potwierdza obserwowaną w latach 1995–2002 tendencję spowolnienia lub nawet zahamowania tempa eutrofizacji jeziora Wigry. Skład gatunkowy Crustacea świadczy o stosunkowo niskiej trofii tego zbiornika, przy czym struktura zooplanktonu może wskazywać na nieco większą podatność na eutrofizację południowej części akwenu. Analizy fizykochemiczne wykazały nieco większą produktywność północnej części Wigier, ale ten rejon już prawie 100 lat temu wykazywał cechy świadczące o wyższej trofii. Obserwuje się tu natomiast kompensację poszczególnych wskaźników TSI i zmniejszenie stężenia biogenów, co wskazuje na stabilizowanie się warunków i zahamowanie tempa eutrofizacji. Południowa część jeziora charakteryzuje się większą podatnością na degradację, a wyróżnikiem podstawowym są większe stężenia biogenów (N, P). Wysoka liczebność zooplanktonu skorupiakowego, z dominacją dużych efektywnych filtratorów, a także zabiegi na rzecz utrzymania wysokiej populacji ryb drapieżnych zapewniają stabilność mezotrofii w większej części jeziora.
Literatura Ber A., 1981: Pojezierze Suwalsko-Augustowskie. Przewodnik geologiczny. Wyd. Geologiczne, Warszawa. Brzęk G., 1988: Stacja hydrobiologiczna na Wigrach, Lublin, Wyd. Lubelskie. Carlson R.E., 1977: A trophic state index for lakes, Limnology and Oceanography, 22: 361–369. Górniak A. i in., 2006: Jeziora Wigierskiego Parku Narodowego – Aktualna jakość i trofia wód, Białystok, Wyd. UwB. Hillbricht-Ilkowska A., Wiśniewski R.J., 1994: Zróżnicowanie troficzne jezior Suwalskiego Parku Krajobrazowego – stan obecny, zmienność wieloletnia, miejsce w klasyfikacji troficznej jezior. Zeszyty Naukowe PAN KN przy Prezydium PAN „Człowiek i Środowisko” 7: 181–200. Karabin A., 1985: Pelagic zooplankton (Rotatoria + Crustacea). Variation in the process of lake eutrophication. I. Structural and quantitative features. Ekol. Pol. 33, 4: 567–616. Karabin A., Ejsmont-Karabin J., 1994: Zespoły zooplanktonu jezior Suwalskiego Parku Krajobrazowego. Zeszyty Naukowe PAN KN przy Prezydium PAN „Człowiek i Środowisko” 7: 215–242. Karabin A., Ejsmont-Karabin J., 1999: Jezioro Wigry – wieloletnia sukcesja zespołów Rotifera i Crustacea. W: B. Zdanowski (red.): Funkcjonowanie i ochrona ekosystemów wodnych na obszarach chronionych. Olsztyn, Wyd. IRŚ: 371–388. Kratzer C.R., Brezonik P.L., 1981: A Carlson-type trophic state index for nitrogen in Florida lakes. Water Res. Bull. 17: 713–715. MacIsaac H.J., Ketelaars H.A.M., Grigorovich I.A., Ramcharan C.W. and Yan N.D., 2000: Modeling Bythotrephes longimanus invasions in the Great Lakes basin based on its European distribution. Arch. Hydrobiol. 149: 1–21. Rudź R., Górniak A., 2005: Zooplankton skorupiakowy jezior harmonijnych Wigierskiego Parku Narodowego. Rocznik Augustowsko-Suwalski 4. Materiały z sesji: 15 lat Wigierskiego Parku Narodowego. Augustowsko-Suwalskie Towarzystwo Naukowe, Suwałki: 105–114. Rybak J.I., 1994: Przegląd słodkowodnych zwierząt bezkręgowych, Biblioteka Monitoringu Środowiska, Warszawa. Zdanowski B., Kamiński M., 1999: Ekosystemy wodne Wigierskiego Parku Narodowego: walory, zagrożenia, ochrona. W: Materiały pokonferencyjne – 10 lat Wigierskiego Parku Narodowego, 14–15 października 1999, Suwałki: 12–20.
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||